Elevated Bioactivity of Ruta graveolens against Cancer Cells and Microbes Using Seaweeds


1
Department of Food Industries, Damietta University, Damietta 34517, Egypt
2
Plant Production Department, College of Food and Agriculture Sciences, King Saud University, P.O. Box 2455, Riyadh 11451, Saudi Arabia
3
Floriculture, Ornamental Horticulture, and Garden Design Department, Faculty of Agriculture (El-Shatby), Alexandria University, Alexandria 21545, Egypt
4
Center for Biodiversity Genomics, University of Guelph, Guelph, ON N1G 1Y2, Canada
5
Precision Agriculture Laboratory, Department of Pomology, Faculty of Agriculture (El-Shatby), Alexandria University, Alexandria 21545, Egypt
*
Author to whom correspondence should be addressed.
Processes 2020, 8(1), 75; https://0-doi-org.brum.beds.ac.uk/10.3390/pr8010075
Submission received: 17 December 2019
/
Revised: 27 December 2019
/
Accepted: 2 January 2020
/
Published: 6 January 2020
(This article belongs to the Special Issue Green Separation and Extraction Processes)
Abstract
:Human cancer and pathogenic microbes cause a significant number of deaths every year. Modulating current sources of natural products that control such diseases becomes essential. Natural algae, such as Ascophyllum nodosum and Ecklonia maxima, can modulate the metabolic processes as well the bioactivities of Ruta graveolens L. The R. graveolens plants were subjected to nine soil drenches of A. nodosum (7 mL L−1), E. maxima (7 mL L−1), or both extracts. Morphological performance, gas exchange parameters, and essential oils (EOs) composition (GC-MS) were studied and the bioactivity was assessed against several cancer cells and pathogenic bacteria or fungi. Treatment with A. nodosum + E. maxima seaweed extracts (SWE) led to the highest morphological performance and gas exchange parameters. The highest antiproliferative, apoptotic, and caspase-3/7 activities of EO were against HeLa in SWE mixture treated plants. The best EO antimicrobial activities were obtained against Staphylococcus aureus and Penicillium ochrochloron. SWE mixtures treated plants showed the best bioactivities against microbes and cancer cells. The highest abundance of 2-undecanone (62%) and 2-nonanone (18%) was found in plants treated with SWE mixtures and caused the best anticancer and antimicrobial effects. Seaweed mixtures act as natural elicitors of pharmaceutical industries and favored 2-undecanone and 2-nonanone in R. graveolens.
1. Introduction
Ruta graveolens (Rutaceae) is an herbaceous species distributed across tropical and temperate areas of the world including South America and Southern Europe and is considered native to the Mediterranean region [1,2]. The plant, with its bitter taste and strong odor, has historical uses in folk medicine of different civilizations [3,4], for example, for the control of menstrual disorders, headache and earache, cramps, and skin inflammation [5]. The leaves contain essential oil (EO) and active ingredients, such as phenols, flavonoids, alkaloids, and saponins [5]. EOs are used in perfumery, pharmaceutical, and food (cheese flavoring) industries. The antioxidant, anti-inflammatory, anthelmintic, antiparasitic, and antimicrobial properties have been described [6]. The main essential oil (EO) compositions of R. graveolens are largely dependent on the ecotype, environment, and external elicitors (for secondary metabolites) [7,8]. Constituents include 2-undecanone, 2-nonanone, and 2-acetoxy tetradecanone [7,8]. External elicitors such as growth regulators and natural biostimulants (e.g., seaweeds) can influence the composition of EO in ornamental medicinal plants and increase their pharmaceutical potential [9,10,11].
Seaweed extract (SWE) consists of natural marine algae, usually collected from seacoasts, then, dried and processed to obtain liquid or dry extracts [12,13,14]. They contain micro and macro elements, i.e., sugars, vitamins, and natural growth regulators that can influence the growth and performance of several horticultural and medicinal crops [15]. Several active compounds have been identified in these SWE such as alginic acid, uronic acid, and phenols in A. nodosum extract and IAA- and GA-like activity in Laminaria extracts [16]. Ecklonia maxima has been successfully used as a biostimulant in several crops and it contains auxins, cytokinins, alginates, amino acids, and sugars [17]. Interest has increased in the use of SWE as a substitute for chemical and synthetic fertilizers, to increase vegetative crop productions, control diseases, enhance flowering and fruiting, and ameliorate stress conditions [9,18]. However, modifying secondary metabolites of medicinal plants has not been studied as extensively. Natural biostimulants, such as SWE can modify the composition and increase the quantity of the secondary metabolite, including EO [9,14]. These modifications can have a great effect against human and plant pathogenic bacteria and fungi, as well as human cancer cells.
Human cancer has caused significant deaths during this century and is considered to be the main cause of deaths worldwide among human diseases [19]. Bacteria and fungi cause many human diseases, leading to death in third world countries. They also influence the production cycle of field crops, including the field growth stage and in post-harvest facilities [20,21]. These microorganisms are usually controlled using antibiotics and commercial reagents; however, microorganisms have developed into multidrug resistant strains, that are able to survive antibiotic or commercial reagent applications [22]. These antibiotics and commercial reagents have several side effects, and therefore a search for safer alternatives is necessary. Accordingly, researchers are mining new resources that can control these bacteria and fungi, such as secondary plant metabolites which include EO. In this respect, SWE mixtures of different algal species, could be a novel approach to increase the medicinal output of the EO of R. graveolens.
The current study investigates the application of two SWEs, namely Ascophyllum nodosum (from Canada) and Ecklonia maxima (from South Africa), on R. graveolens secondary metabolites. This study proposed that these natural biostimulants are modulating the growth and EO constituents, leading to enhanced bioactivity against selected cancer cells, bacteria, and fungi. Indeed, these effects have potential for future applications in pharmaceutical and agricultural industries. This is the first study exploring the use of marine algal species mixtures as elicitors of main EO constituents of the aromatic R. graveolens.
2. Results
2.1. Morphological Responses to SWE
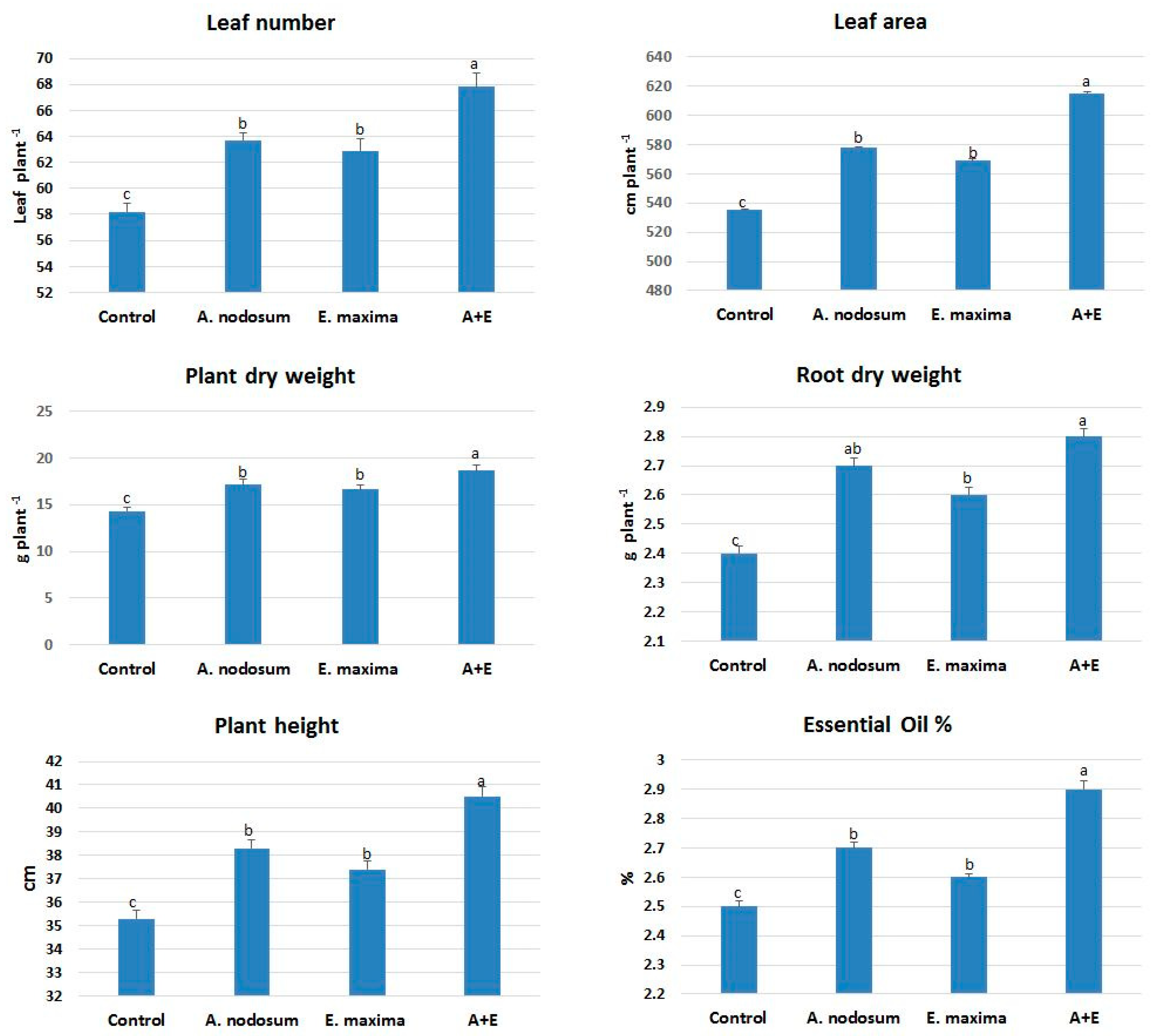
Morphological responses of the plant are the first indicators of the metabolic changes in response to external elicitors. Leaf numbers increased following treatment with A. nodosum, E. maxima and their combination (A + E) as compared with a control (Figure 1). The A + E treatment at 7 mL/L increased leaf number to 67.8 ± 1.1, which is the highest value as compared with other treatments. The leaf area increased in plants treated with A + E as compared with other treatments. Plant root dry weights also increased in plants subjected to SWE. In addition, there was an increase in those morphological parameters associated with increases in plant height. The mean plant height increased from 35.3 cm (control) to 37.4 cm (E. maxima). The EO ratio showed increases in the SWE treated plants.
2.2. Gas Exchange and SWE Mixture
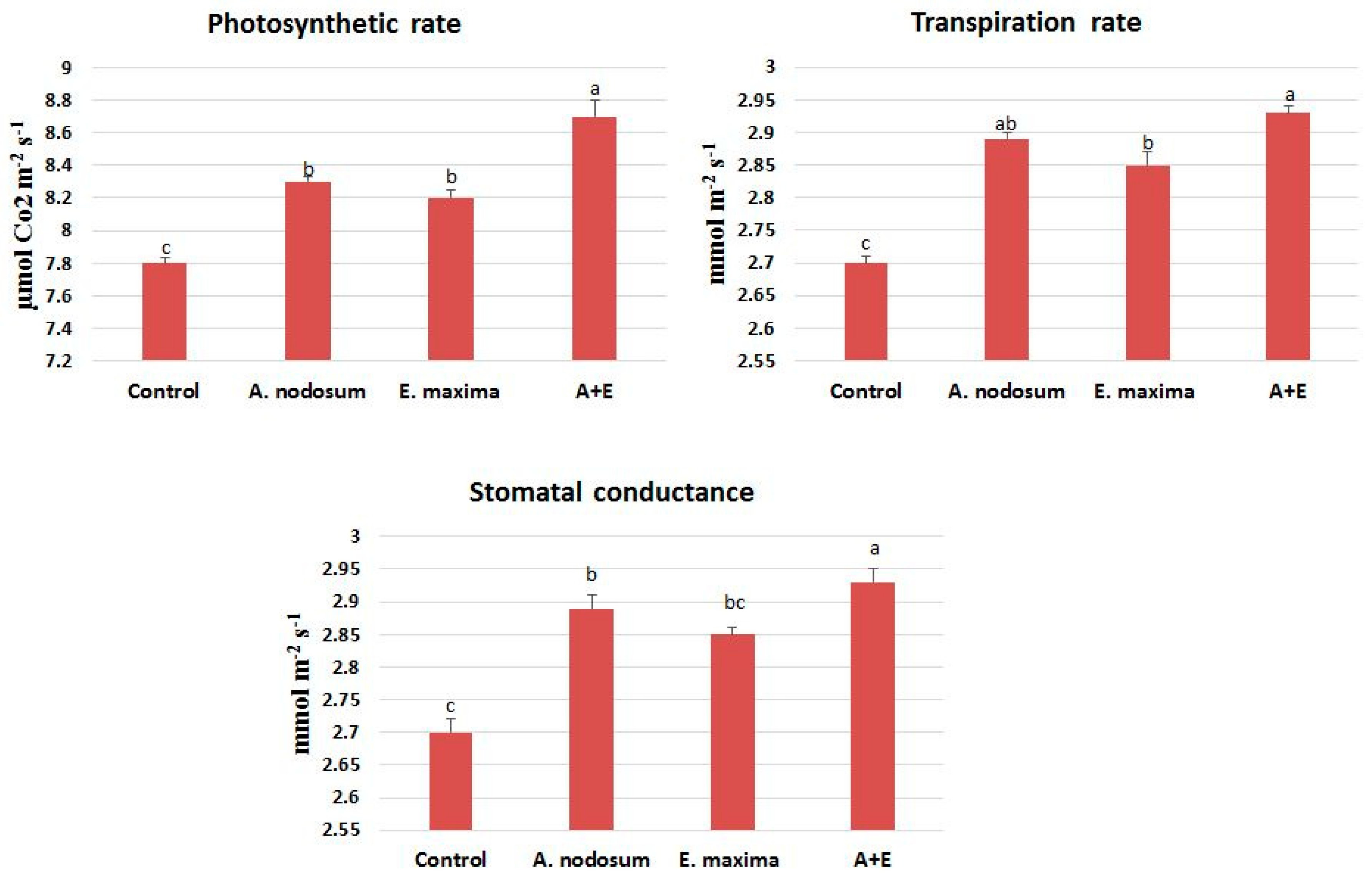
In general, gas exchange parameters are strongly associated with morphological and physiological performance of a plant. The A increased in R. graveolens treated with different SWE, as shown in Figure 2. The highest peak in the mean photosynthetic rate was in response to the A + E mixture treatment as compared with A. nodosum or E. Maxima SWE only. Indeed, the control showed much lower values of A as compared with the SWE treatments. E rates were higher in plants treated with SWE mixture and the highest value was 2.93 ± 0.1 mmol m−2 s−1. The stomatal conductance (gs) was highest (127.6 ± 1.4 mmol m−2 s−1) in plants treated with the SWE mixture as compared with that of single SWE and the control.
2.3. Essential Oil and SWE Mixture
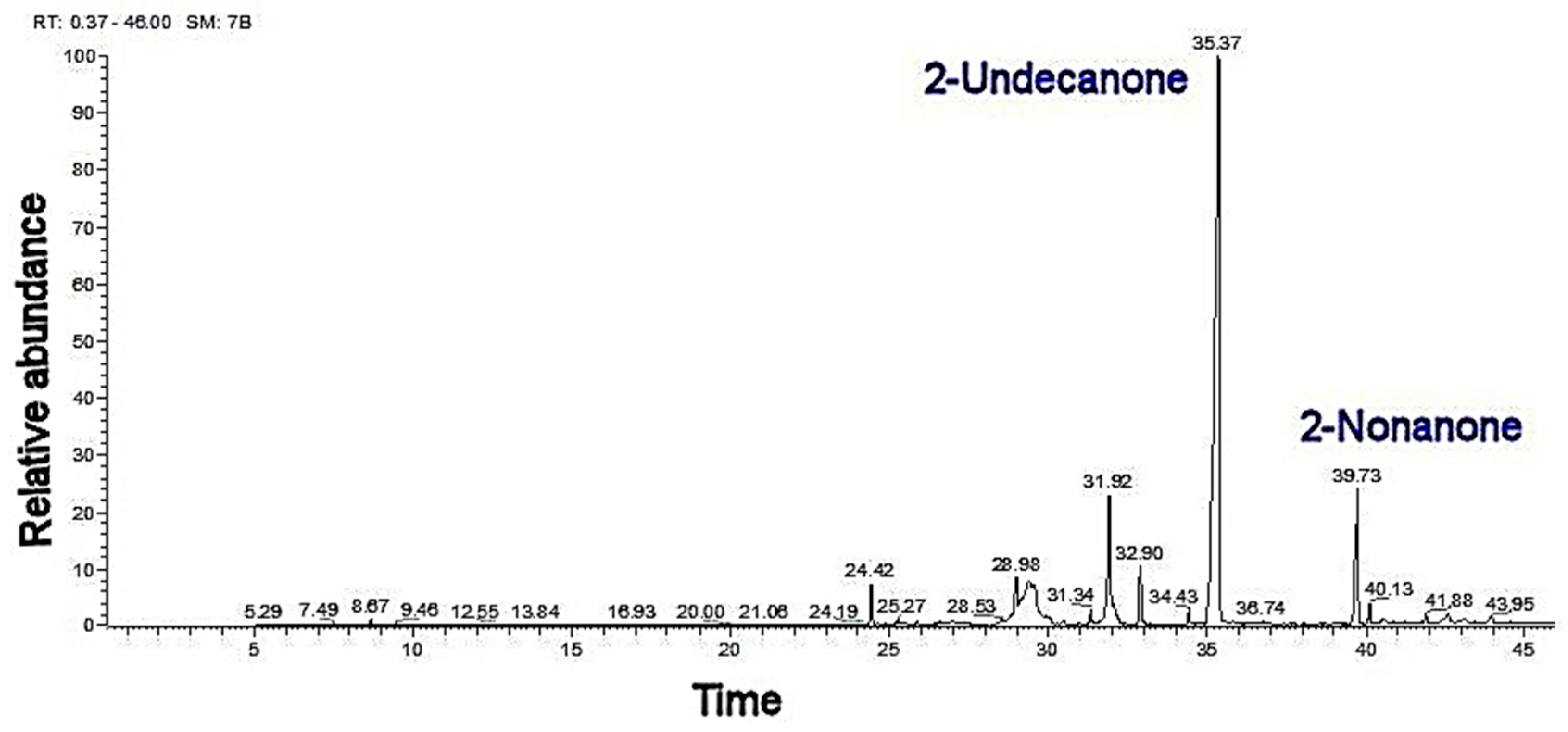
Essential oils are the major secondary metabolite of these plants which are very sensitive to external elicitors. The EO was mainly composed of 2-undecanone (62%) and 2-nonanone (18.01%) as shown in Table 1 and Figure 3. A. nodosum SWE showed higher increases in major essential oil constituents as compared with that of the control plants. The application of SWE mixtures showed the highest increases in major essential oil constituents. The EO of R. graveolens was affected by SWE and their mixtures. The SWE mineral analyses showed moderate percentages of nitrogen (N, 0.5% and 0.2%) and phosphorus (P2O5, 0.19% and 0.14%) and potassium (K2O, 0.7% and 0.2%) in A. nodosum and E. maxima, respectively. Other important minerals were found such as magnesium (0.1% and 0.1%) and calcium (0.9% and 1.0%) in A. nodosum and E. maxima, respectively. Trace ratios (1−5% × 10−4%) of heavy metals (iron, zinc, manganese, boron, and copper) were detected.
2.4. Antiproliferative Activities and Apoptotic Assay
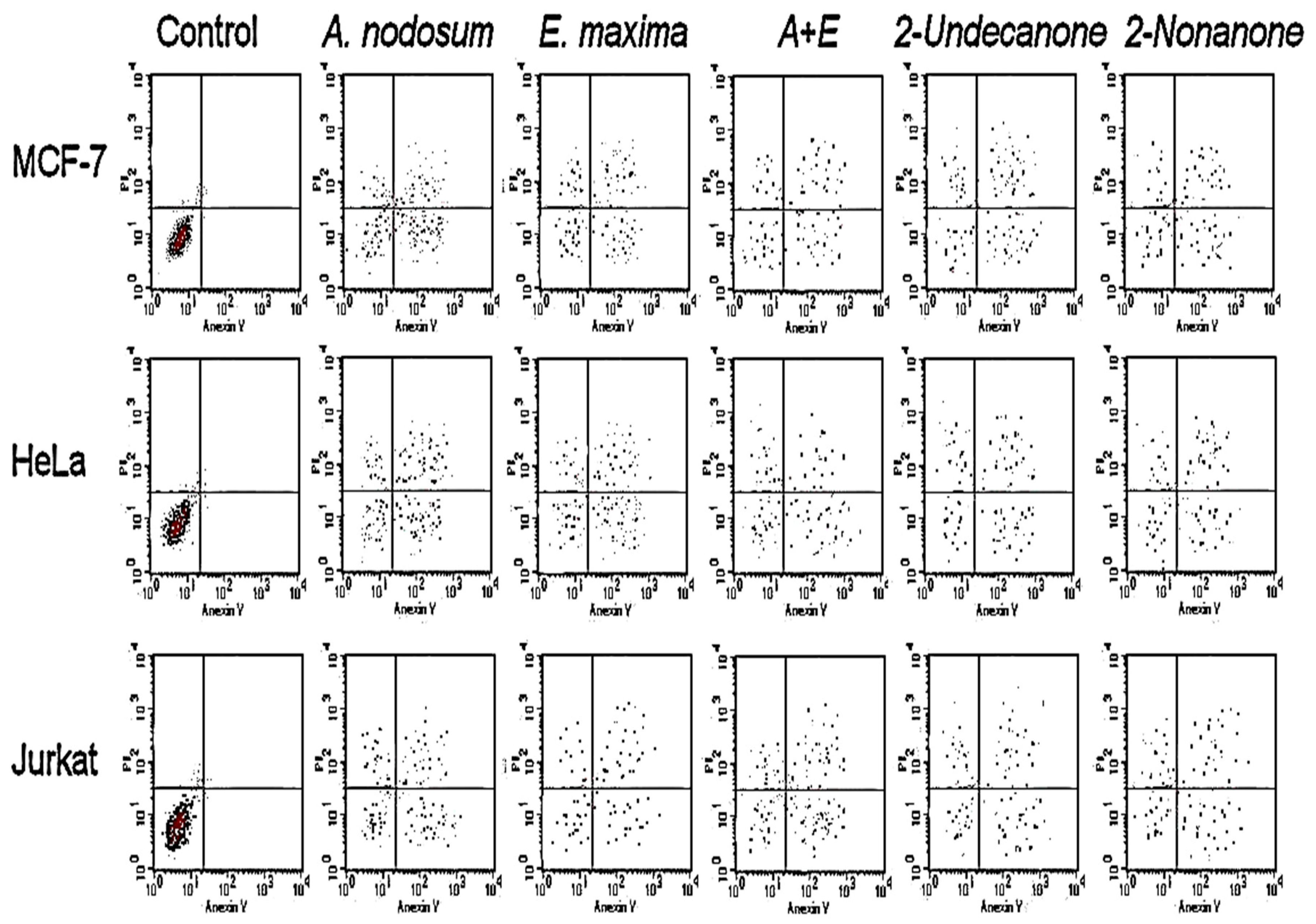
The antiproliferative activity of Eos, IC50 (µg mL−1), as affected by SWE treatments, is presented in Table 2. The EO of control plants showed the lowest antiproliferative activity against different cancer cell lines. The EO of plants subjected to SWE treatments with E. maxima/A. nodosum were stronger against cancer cell lines than that of the control plants. The A + E treated plants EO showed the highest antiproliferative activities expressed as IC50 (8.11, 4.3, 17.05, 82.2, and 70.53 µg mL−1) for MCF-7, HeLa, JURKAT, T24, and HT29. All EO, 2-undecanone, and 2-nonanone, had no antiproliferative activity against normal cancer cell lines of HEK-293. The apoptotic assay (Figure 4) revealed increased accumulation of necrotic cells, as well as early and late apoptotic cells in all treatments except in the control.
2.5. Caspase-3/7 Activity Study
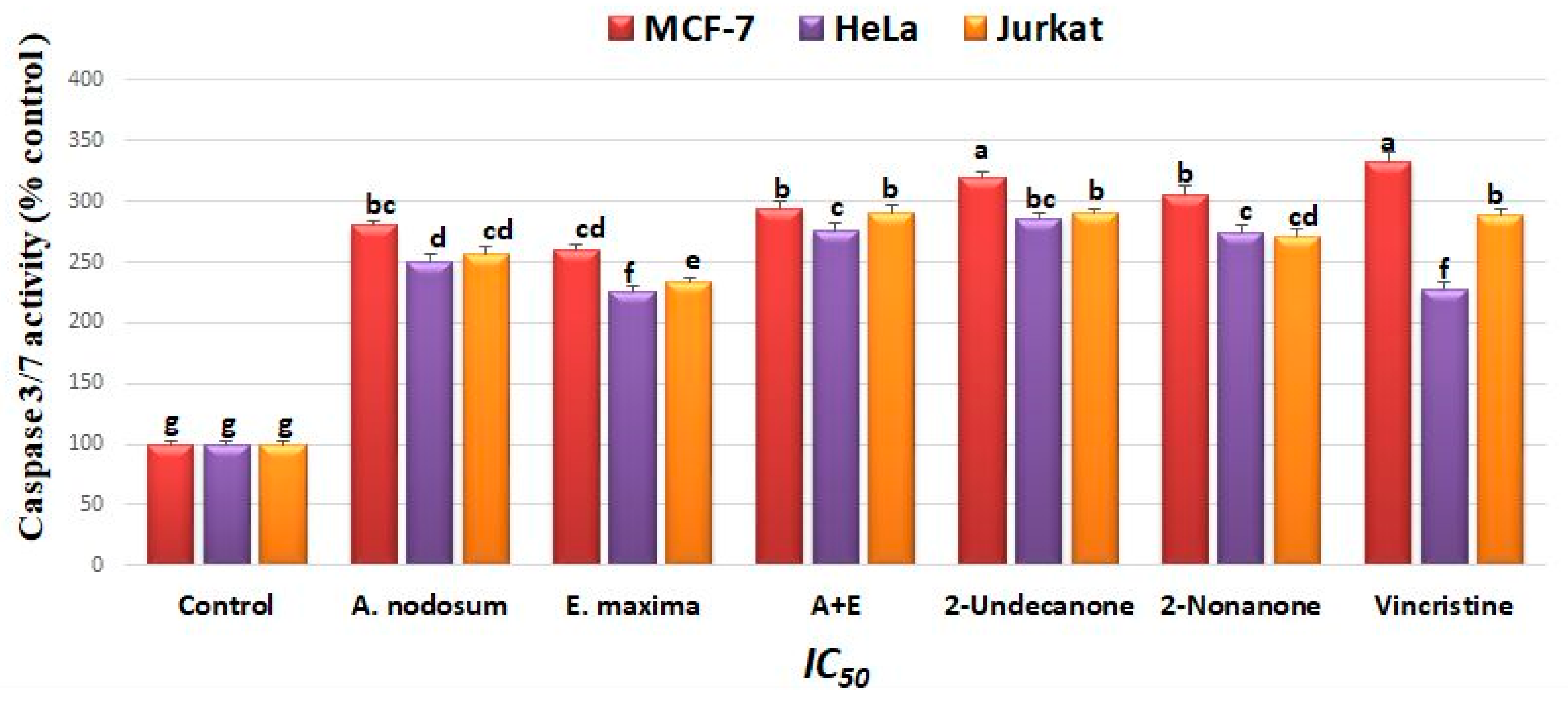
The effects of EO, 2-undecanone, and 2-nonanone on caspase-3/7 activity in cancer cells are shown in Figure 5. The activity of caspase 3/7 increased in all seaweed treated plants and the mixture of seaweeds was higher than individual ones as well as control cells. Treatments with 2-undecanone and 2-nonanone showed higher activity in cancer cell lines than other treatments.
2.6. EO and Antibacterial Activities
The antibacterial assay results are presented in Table 3. The MIC ranged from 0.04 to 0.42 mg mL−1 and MBC from 0.08 to 0.93 mg mL−1. Control plant EOs exhibited low antibacterial potential against most bacteria, whereas EO of plants subjected to SWE with E. maxima/A. nodosum were more active against most bacteria. The A + E treated plants EO were the most active (as compared with other extracts) showing high MIC (0.06 to 0.29 mg mL−1) and MBC (0.13 to 0.71 mg mL−1), S. aureus and P. c. subsp.
2.7. EO and Antifungal Activities
The EO, 2-undecanone and 2-nonanone bioactivities against fungi were determined, as shown in Table 4. High MIC and MFC values were measured in EO obtained from SWE treated plants. The values ranged from 0.06 to 0.32 mg mL−1 and from 0.10 to 0.73 g L−1 for MIC and MFC, respectively. EOs obtained from A + E treatments were the most active showing low MIC and MFC. The most sensitive fungi for A + E treatment was P. ochrochloron. The activities of Eos, as well as the 2-undecanone and 2-nonanone, were comparable to commercial agents.
3. Discussion
The effects of SWE on the morphological characteristics of plants have been documented in several previous studies [15,23]. The stimulatory effects of SWE is greatly attributed to macro- and microelements, possible available natural growth regulators (cytokinins and auxins), oligosaccharides, and vitamins. These nutrients and growth regulators stimulate the cellular metabolism and the morphological growth. The nutrients found in SWE used here include nitrogen, phosphorus, potassium, magnesium, calcium, and micro elements, all of which enhance the growth of plants.
The increases in the morphological performance of treated plants is strongly associated with gas exchange, as found in several studies [9,10,14,24]. There were increases in A in R. graveolens, treated with different SWE. These increases are strongly associated with improved metabolic performance in seaweed treated plants. A previous investigation suggested that an increased leaf chlorophyll content in SWE treated plants [25] leads to increase A, followed by increased gs and E [10]. In addition, it was suggested that seaweed can reduce the chlorophyll degradation in treated plants [26]. The SWE mixture (A + E) resulted in the best morphological and physiological performance in plants, which is strongly related to the nutritional composition of the two seaweeds. This nutrient mixture showed the best results and no previous study has found comparable results.
The control R. graveolens plants showed an EO profile matching global and local standards [7,27]. There were major shifts in EO dominant compounds of R. graveolens plants subjected to SWE mixtures. These shifts in the EO profiles related to SWE treatment and peaked in SWE mixture treatment. Specifically, major EO constituents increased (2-undecanone and 2-nonanone). These constituents (ketones) are highly favored in the food industry (e.g., flavoring cheese) and in the perfumery industry as well [27]. High medicinal value is associated with industrial and pharmaceutical potential, as we found. Studies on mint and basil (medicinal plants) reported similar effects of single SWE on EO ratio and composition [11]. However, the use of SWE mixtures to increase secondary metabolites (specific EO constituents) is a novel methodology, resulting in synergism by both SWEs.
To study the effects SWE as external elicitors that can elevate the biological activities of R. graveolens, the biological activities against cancer cells were investigated. In this study, there were obvious antiproliferative activities of R. graveolens EO, 2-undecanone, and 2-nonanone against specific cancer cells. In addition, the SWE mixture treated plants showed the highest antiproliferative activity. Leaf extracts of R. graveolens had anticancer activities against breast, colon, and prostate cancers [28]. In addition, specific alkaloids in the leaves were associated with anticancer activities against MCF-7 and others [29]. However, studies regarding the anticancer potential of EO are lacking. The increased activities of caspase-3 and -7 following treatment with seaweed treated plants EOS, 2-undecanone, and 2-nonanone are strong indicators of apoptosis by DNA fragmentation [30,31,32].
To further investigate the elevated biological activities of EO against different microorganisms, an antimicrobial study was conducted. The EO composition shifts by SWE mixtures maximized the antibacterial and antifungal potential, as shown before. These shifts included increases of 2-undecanone and 2-nonanone. The 2-undecanone had almost 50% of the antibacterial and antifungal activities values of EO obtained from plants subjected to SWE mixture which is related to the ratio of the 2-undecanone in the EO of these plants. Other reports indicated obvious potential of undecanone and nonanone for microbial control [7,33]. The 2-undecano found in Indian EO of R. graveolens was related to the control of S. aureus growth using the inhibition zone method [7]. The bioactivity of the EO of Houttuynia cordata Thunb, (medicinal plant) is caused by 2-undecanone [34]. South African R. graveolens EO had comparable ratios of nonanone and undecanone and activities against B. cereus and S. aureus [35].
This study explored the elevated bioactivities in response to novel external elicitor mixture to two SWE. Furthermore, it associated the EO main constituents (2-undecanone and 2-nonanone) and bioactivities against cancer cells. Modulation of R. graveolens EO by SWE mixture, showed clear anticancer, antibacterial, and antifungal potential leading to enriching natural sources of pathogens control, in addition to providing promising potential as anticancer agents. Plant pathogens studied here have great negative effects on crop production and post-harvest. The SWE mixture modulated EO found here have great potential as safe alternative to bactericides and fungicides, against plant pathogens.
4. Materials and Methods
4.1. Plant Material, Treatments, and Microorganisms
R. graveolens L. uniform plants were obtained (March 2017 and 2018, in two successive seasons), then identified and vouchered by Hosam Elansary and registered at the Faculty of Agriculture, Alexandria University (voucher no. 002314-Elansary). Plants were raised in 2.1 L pots containing brown peat and perlite (3:1 w/w) and supplemented with Crystalon® (20% N: 20% P: 20% K, 2 gL−1 media) under controlled environmental conditions. The temperature ranged between 15.2 °C (night) and 27.6 °C (day); relative humidity between 68% and 71%; photosynthetically active radiation (PAR) around 1000 m−2 μmol m−2 s−1 at 12.00 p.m.; and daily watering of 38 to 50 mL plant −1.
The plants were subjected to 9 treatments of SWE (Ascophyllum nodosum, Stella Maris TM, Acadian Seaplants, Guelph, ON, Canada; 7 mL L−1 with irrigation water), SWE (Ecklonia maxima, Kelpak TM, South Africa; 7 mL/L with irrigation water), or a mixture of both SWEs (1:1, 7 mL L−1 with irrigation water). Treatments were repeated every 6 d until achieving 9 treatments. Untreated plants were considered as a control. Plants were grouped into three blocks/repetitions (n = 3) containing 10 replicates per treatment for a total of 120 plants, in a factorial experimental design. The fungi and bacteria were obtained from King Saud University. The 2-undecanone, 2-nonanone, and HPLC-grade chemicals were obtained from Sigma-Aldrich, Berlin, Germany.
4.2. Measurements
After the 9 treatments, plant heights (cm) were determined before being harvested, and the roots were cleaned for further measurements. Leaf number and leaf area were also determined. Leaf area was determined by digital area meter. Plants were air dried in an oven dryer at 35 °C (to maintain the oil composition and ratio) and the total dry weight and root dry weight were measured. The stomatal conductance (gs), net photosynthetic rate (A), and transpiration rate (E) were determined using a portable photosynthesis system analyzer (ADC BioScientific, LCi, Bioscientific Ltd., Hoddesdon, UK) on fully expanded leaves, in clear, sunny conditions [36].
4.3. Essential Oil Isolation, Gas Chromatography/Mass Spectrometry (GC-MS), and Analyses of Seaweeds
The dried leaves were ground and hydro-distilled for 1 h using a Clevenger apparatus and the EO were dried over anhydrous sodium sulfate and stored at 4 °C. The EO were subjected to gas chromatography (Hewlett Packard 5890) and compounds identification were as described before [11,37,38]. Briefly, separation was accomplished with a TG-1MS narrow bore column (length 30 m × 0.32 mm i.d. and 0.25 μm film thickness). Helium was used as the carrier gas. The oven temperature was programmed to increase from an initial temperature of 45° to 165 °C, at 4 °C min−1, with a holding time of 2 min at 45 °C, then, 15 °C min−1 to 280 °C, with a final holding time of 15 min. A total of 2 μL of each sample was injected at 250 °C on a splitless mode flow (1 mL min−1), and splitless time (3 min), then, split flow (10 mL min−1). Reference literature was also used to confirm compounds [7] as well as chemical standards (Sigma-Aldrich, Berlin, Germany). The mineral profile of seaweeds used in this study were determined using inductively coupled plasma spectroscopic analysis (ICPSA) (Optima 4300DV, Perkin-Elmer, Waltham, MA, USA) and ICP-OES as described before [39].
4.4. Antiproliferative Activity and Apoptotic Cell Population
The antiproliferative activities of the EO were tested against several cancer cell lines, including MCF-7, HeLa, Jurkat, HT-29, and T24, as well as normal cells of HEK-293 following [40]. Five doses of the EO, 2-undecanone and 2-nonanone were used to reach a final concentration of 50, 100, 200, 300, and 400 µg/mL culture media. Vinblastine sulfate and taxol were used as positive controls. The antiproliferative activities were expressed as the IC50 (µg mL−1) values of the EO, by plotting the percentage of cell viability against the extract concentration. The IC50 was used to measure the apoptotic cell population using flow cytometry (FAC Scan, Becton Dickinson, Burlington, MA, USA) following [31,41].
4.5. Caspase-Glo 3/7 Assay
The effects of EO on caspase-3/7 activity against cancer cell lines was investigated using Caspase-Glo 3/7 Assay kit (Promega, Berlin, Germany). Twenty-four hour cultured cell lines (Roswell Park Memorial Institute (RPMI) growth medium, Sigma-Aldrich) in IC50 of EOs, 2-undecanone and 2-nonanone or DMSO (solvent control) were used [31]. Then, 100 µL of caspase-Glo 3/7 reagent was added to 96-well plates containing growing media, mixed, and then incubated for 1 h at room temperature. An Infinite M2000 Pro™ (Tecan, Vienna, Austria) was used to detect the luminescence and the activity was expressed as a percentage of the untreated samples.
4.6. Antibacterial Activities
The bacteria used were Bacillus cereus (ATCC 14579), Staphylococcus aureus (ATCC 6538), Micrococcus flavus (ATCC 10240), and Listeria monocytogenes (clinical isolate) as Gram-positive bacteria and Escherichia coli (ATCC 35210), Dickeya solani (D s0432-1), Pseudomonas aeruginosa (ATCC 27853), Pectobacterium carotovorum subsp. carotovorum (ATCC 15713), and Pectobacterium atrosepticum (ATCC 33260) as Gram-negative bacteria. The microdilution method [31,41,42] was used to calculate the minimum inhibitory concentration (MIC) and minimum bactericidal (MBC) concentration. Microtiter plates (96-well) containing a serial concentration of the essential oil in each well mixed with bacterial inoculum (1.0 × 104 CFU per well) in 100 μL tryptic soy broth were incubated at 37 °C for 24 h in a rotary shaker. The minimum inhibitory concentration (MIC) was defined as the lowest concentration of essential oil that exhibited no visible growth using a binocular microscope and was determined following the incubation period of the microtiter plates. The minimum bactericide concentration (MBC), which was defined as the lowest concentration that caused no visible growth and indicated the killing of 99.5% of the inoculum, was determined using serial subculturing of the essential oil (2 μL). The optical density was determined at a wavelength of 655 nm. A positive control was used (streptomycin, 0.01 to 10 mg/mL), as well as a negative one (DMSO, 1%).
4.7. Antifungal Activities
Six fungi were used, including Aspergillus niger (ATCC 6275), Penicillium funiculosum (ATCC 56755), Aspergillus flavus (ATCC 9643), Aspergillus ochraceus (ATCC 12066), Penicillium ochrochloron (ATCC 48663), and Candida albicans (ATCC 12066). The minimum inhibitory (MIC) and minimum fungicidal (MFC) concentrations of the microdilution method were used [41,42,43,44,45].
4.8. Statistical Analyses
Data obtained from the two seasons of 2017 and 2018 were subjected to analyses of variance (ANOVA) in SPSS v. 22 and showed no significant differences between the two seasons. Data from both seasons were pooled together and the least significant differences was used to compare all means.
5. Conclusions
This is the first study to explore the use of marine algal species mixtures as elicitors of main EO constituents of the aromatic R. graveolens, an application which could be beneficial to the pharmaceutical industry as antimicrobial and anticancer sources. The application of A. nodosum + E. maxima SWE led to superior morphological performance and maximum gas exchange values in treated plants. The main EO constituents were 2-undecanone, 2-nonanone, 2-acetoxy tetradecane, and nonyl cyclopropanecarboxylate. The highest ratios of 2-undecanone, 2-nonanone, and nonyl cyclopropanecarboxylate were detected in plants treated with A. nodosum + E. maxima SWE. Morphological and physiological parameters, as well as EO main constituents associated with the bioactivity of the EO of treated plants against bacteria, fungi, and cancer cells. The highest antibacterial and antifungal activities were found against S. aureus and P. ochrochloron, respectively. The highest antiproliferative and necrotic activities were against HeLa MCF-7 and Jurkat. The anticancer activities were confirmed by increased activities of the caspase-3 and -7 enzymes in treated cancer cells. The SWE mixtures are recommended as natural elicitors of 2-undecanone and 2-nonanone of R. graveolens EO which have good potential in the agricultural and pharmaceutical industries. Further investigations could be conducted in this application on different crops.
Author Contributions
Conceptualization, E.A.M., H.O.E., and F.A.A.-M.; data curation, E.A.M., H.O.E., and D.O.E.-A.; formal analysis, E.A.M., H.O.E., D.O.E.-A., and F.A.A.-M.; funding acquisition, F.A.A.-M.; project administration, E.A.M.; software, E.A.M. and F.A.A.-M.; validation, H.O.E. and D.O.E.-A.; visualization, E.A.M., H.O.E., and D.O.E.-A.; writing—original draft, H.O.E.; writing—review and editing, E.A.M. and H.O.E. All authors have read and agreed to the published version of the manuscript.
Funding
The study was funded by the King Saud University through Researchers Supporting Project number (RSP-2019/118).
Acknowledgments
The study was funded by the King Saud University through Researchers Supporting Project number (RSP-2019/118).
Conflicts of Interest
Authors declare no conflict of interest.
References
- Ueng, Y.-F.; Chen, C.-C.; Huang, Y.-L.; Lee, I.J.; Yun, C.-H.; Chen, Y.-H.; Huang, C.-C. Effects of aqueous extract of Ruta graveolens and its ingredients on cytochrome P450, uridine diphosphate (UDP)-glucuronosyltransferase, and reduced nicotinamide adenine dinucleotide (phosphate) (NAD(P)H)-quinone oxidoreductase in mice. J. Food Drug Anal. 2015, 23, 516–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, F.G.E.; Mendes, F.R.D.S.; Assunção, J.C.D.C.; Maria Pinheiro Santiago, G.; Aislania Xavier Bezerra, M.; Barbosa, F.G.; Mafezoli, J.; Rodrigues Rocha, R. Seasonal variation, larvicidal and nematicidal activities of the lef essential oil of Ruta graveolens L. J. Essent. Oil Res. 2014, 26, 204–209. [Google Scholar] [CrossRef]
- Pollio, A.; De Natale, A.; Appetiti, E.; Aliotta, G.; Touwaide, A. Continuity and change in the Mediterranean medical tradition: Ruta spp. (rutaceae) in Hippocratic medicine and present practices. J. Ethnopharmacol. 2008, 116, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Dob, T.; Dahmane, D.; Gauriat-Desrdy, B.; Daligault, V. Volatile Constituents of the Essential Oil of Ruta chalepensis L. subsp. Angustifolia (Pers.) P. Cout. J. Essent. Oil Res. 2008, 20, 306–309. [Google Scholar] [CrossRef]
- Mejri, J.; Abderrabba, M.; Mejri, M. Chemical composition of the essential oil of Ruta chalepensis L: Influence of drying, hydro-distillation duration and plant parts. Ind. Crop. Prod. 2010, 32, 671–673. [Google Scholar] [CrossRef]
- Amdouni, T.; Ben Abdallah, S.; Msilini, N.; Merck, F.; Chebbi, M.; Lachaal, M.; Karray-Bouraoui, N.; Ouerghi, Z.; Fernandez, X. Effect of salt stress on the antimicrobial activity of Ruta chalepensis essential oils. Acta Physiol. Plant. 2016, 38, 147. [Google Scholar] [CrossRef]
- Reddy, D.N.; Al-Rajab, A.J. Chemical composition, antibacterial and antifungal activities of Ruta graveolens L. volatile oils. Cogent Chem. 2016, 2, 1220055. [Google Scholar] [CrossRef]
- Yaacob, K.B.; Abdullah, C.M.; Joulain, D. Essential Oil of Ruta graveolens L. J. Essent. Oil Res. 1989, 1, 203–207. [Google Scholar] [CrossRef]
- Al-Ghamdi, A.A.; Elansary, H.O. Synergetic effects of 5-aminolevulinic acid and Ascophyllum nodosum seaweed extracts on Asparagus phenolics and stress related genes under saline irrigation. Plant Physiol. Bioch. 2018, 129, 273–284. [Google Scholar] [CrossRef]
- Elansary, H.O. Green roof Petunia, Ageratum, and Mentha responses to water stress, seaweeds, and trinexapac-ethyl treatments. Acta Physiol. Plant. 2017, 39, 145. [Google Scholar] [CrossRef]
- Elansary, H.O.; Yessoufou, K.; Shokralla, S.; Mahmoud, E.A.; Skaicka-Wozniak, K. Enhancing mint and basil oil composition and antibacterial activity using seaweed extracts. Ind. Crops Prod. 2016, 92, 50–56. [Google Scholar] [CrossRef]
- Cheong, K.-L.; Qiu, H.-M.; Du, H.; Liu, Y.; Khan, M.B. Oligosaccharides Derived from Red Seaweed: Production, Properties, and Potential Health and Cosmetic Applications. Molecules 2018, 23, 2451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, S.; Wang, R.; Zhang, M.; Meng, X.; Liu, H.; Wang, L. Insights into the Fouling Propensities of Natural Derived Alginate Blocks during the Microfiltration Process. Processes 2019, 7, 858. [Google Scholar] [CrossRef] [Green Version]
- Elansary, H.O.; Skalicka-Woźniake, K.; King, I.W. Enhancing stress growth traits as well as phytochemical and antioxidant contents of Spiraea and Pittosporum under seaweed extract treatments. Plant Physiol. Biochem. 2016, 105, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Elansary, H.O.; Yessoufou, K.; Abdel-Hamid, A.M.E.; El-Esawi, M.A.; Ali, H.M.; Elshikh, M.S. Seaweed Extracts Enhance Salam Turfgrass Performance during Prolonged Irrigation Intervals and Saline Shock. Front. Plant Sci. 2017, 8, 830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ertani, A.; Francioso, O.; Tinti, A.; Schiavon, M.; Pizzeghello, D.; Nardi, S. Evaluation of Seaweed Extracts from Laminaria and Ascophyllum nodosum spp. as Biostimulants in Zea mays L. Using a Combination of Chemical, Biochemical and Morphological Approaches. Front. Plant Sci. 2018, 9, 428. [Google Scholar] [CrossRef] [PubMed]
- Kocira, S.; Szparaga, A.; Kuboń, M.; Czerwińska, E.; Piskier, T. Morphological and Biochemical Responses of Glycine max (L.) Merr. to the Use of Seaweed Extract. Agronomy 2019, 9, 93. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Giordano, M.; Cardarelli, M.; Cozzolino, E.; Mori, M.; Kyriacou, C.M.; Bonini, P.; Colla, G. Plant- and Seaweed-Based Extracts Increase Yield but Differentially Modulate Nutritional Quality of Greenhouse Spinach through Biostimulant Action. Agronomy 2018, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Zaorsky, N.G.; Churilla, T.M.; Egleston, B.L.; Fisher, S.G.; Ridge, J.A.; Horwitz, E.M.; Meyer, J.E. Causes of death among cancer patients. Ann. Oncol. 2017, 28, 400–407. [Google Scholar] [CrossRef]
- Sharma, K.; Mahato, N.; Lee, Y.R. Systematic study on active compounds as antibacterial and antibiofilm agent in aging onions. J. Food Drug Anal. 2018, 26, 518–528. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Zheng, Y.; Xiang, F.; Li, S.; Yang, G. Antifungal activity of Momordica charantia seed extracts toward the pathogenic fungus Fusarium solani L. J. Food Drug Anal. 2016, 24, 881–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Duin, D.; Paterson, D.L. Multidrug-Resistant Bacteria in the Community: Trends and Lessons Learned. Infect. Dis. Clin. 2016, 30, 377–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arioli, T.; Mattner, S.W.; Winberg, P.C. Applications of seaweed extracts in Australian agriculture: Past, present and future. J. Appl. Phycol. 2015, 27, 2007–2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, C.S.; Araujo, L.; Alves Chaves, J.A.; DaMatta, F.M.; Rodrigues, F.A. Water relation, leaf gas exchange and chlorophyll a fluorescence imaging of soybean leaves infected with Colletotrichum truncatum. Plant Physiol. Biochem. 2018, 127, 119–128. [Google Scholar] [CrossRef]
- Blunden, G.; Jenkins, T.; Liu, Y.-W. Enhanced leaf chlorophyll levels in plants treated with seaweed extract. J. Appl. Phycol. 1996, 8, 535–543. [Google Scholar] [CrossRef]
- Whapham, C.A.; Blunden, G.; Jenkins, T.; Hankins, S.D. Significance of Betaines in the Increased Chlorophyll Content of Plants Treated with Seaweed Extract. J. Appl. Phycol. 1993, 5, 231–234. [Google Scholar] [CrossRef]
- Attia, E.Z.; Abd El-Baky, R.M.; Desoukey, S.Y.; El Hakeem Mohamed, M.A.; Bishr, M.M.; Kamel, M.S. Chemical composition and antimicrobial activities of essential oils of Ruta graveolens plants treated with salicylic acid under drought stress conditions. Future J. Pharm. Sci. 2018, 4, 254–264. [Google Scholar] [CrossRef]
- Fadlalla, K.; Watson, A.; Yehualaeshet, T.; Turner, T.; Samuel, T. Ruta graveolens extract induces DNA damage pathways and blocks Akt activation to inhibit cancer cell proliferation and survival. Anticancer Res. 2011, 31, 233–241. [Google Scholar]
- Schelz, Z.; Ocsovszki, I.; Bozsity, N.; Hohmann, J.; Zupko, I. Antiproliferative Effects of Various Furanoacridones Isolated from Ruta graveolens on Human Breast Cancer Cell Lines. Anticancer Res. 2016, 36, 2751–2758. [Google Scholar]
- Alvarado-Sansininea, J.J.; Sanchez-Sanchez, L.; Lopez-Munoz, H.; Escobar, M.L.; Flores-Guzman, F.; Tavera-Hernandez, R.; Jimenez-Estrada, M. Quercetagetin and Patuletin: Antiproliferative, Necrotic and Apoptotic Activity in Tumor Cell Lines. Molecules 2018, 23, 2579. [Google Scholar] [CrossRef] [Green Version]
- Elansary, H.O.; Szopa, A.; Kubica, P.; Al-Mana, F.A.; Mahmoud, E.A.; El-Abedin, T.K.A.Z.; Mattar, M.A.; Ekiert, H. Phenolic Compounds of Catalpa speciosa, Taxus cuspidata, and Magnolia acuminata have Antioxidant and Anticancer Activity. Molecules 2019, 24, 412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, R.A.V.; Megeney, L.A. Evolution of caspase-mediated cell death and differentiation: Twins separated at birth. Cell Death Differ. 2017, 24, 1359–1368. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.S.; Joshi, N.; Padalia, R.C.; Singh, V.R.; Goswami, P.; Kumar, A.; Iqbal, H.; Verma, R.K.; Chanda, D.; Chauhan, A.; et al. Chemical Composition and Allelopathic, Antibacterial, Antifungal, and Antiacetylcholinesterase Activity of Fish-mint (Houttuynia cordataThunb.) from India. Chem. Biodivers. 2017, 14, e1700189. [Google Scholar] [CrossRef] [PubMed]
- Viljoen, A.M.; Moolia, A.; van Vuuren, S.F.; van Zyl, R.L.; Başer, K.H.C.; Demirci, B.; Özek, T.; Trinder-Smith, T.H. The Biological Activity and Essential Oil Composition of 17 Agathosma (Rutaceae) Species. J. Essent. Oil Res. 2006, 18 (Suppl. 1), 2–16. [Google Scholar] [CrossRef]
- França Orlanda, J.F.; Nascimento, A.R. Chemical composition and antibacterial activity of Ruta graveolens L. (Rutaceae) volatile oils, from São Luís, Maranhão, Brazil. S. Afr. J. Bot. 2015, 99, 103–106. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Elansary, H.O.; El-Shanhorey, N.A.; Abdel-Hamid, A.M.E.; Ali, H.M.; Elshikh, M.S. Salicylic Acid-Regulated Antioxidant Mechanisms and Gene Expression Enhance Rosemary Performance under Saline Conditions. Front. Physiol. 2017, 8, 716. [Google Scholar] [CrossRef]
- Elansary, H.O.; Ashmawy, N.A. Essential Oils of Mint between Benefits and Hazards. J. Essent. Oil Bear Plants 2013, 16, 429–438. [Google Scholar] [CrossRef]
- Elansary, H.O.; Mahmoud, E.A. Basil cultivar identification using chemotyping still favored over genotyping using core barcodes and possible resources of antioxidants. J. Essent. Oil Res. 2015, 27, 82–87. [Google Scholar] [CrossRef]
- Elansary, H.O.; Norrie, J.; Ali, H.M.; Salem, M.Z.M.; Mahmoud, E.A.; Yessoufou, K. Enhancement of Calibrachoa growth, secondary metabolites and bioactivity using seaweed extracts. BMC Complement. Altern. Med. 2016, 16, 341. [Google Scholar] [CrossRef] [Green Version]
- Parry, J.; Su, L.; Moore, J.; Cheng, Z.; Luther, M.; Rao, J.N.; Wang, J.Y.; Yu, L.L. Chemical compositions, antioxidant capacities, and antiproliferative activities of selected fruit seed flours. J. Agric. Food Chem. 2006, 54, 3773–3778. [Google Scholar] [CrossRef]
- Elansary, H.O.; Szopa, A.; Kubica, P.; Ekiert, H.; Mattar, M.A.; Al-Yafrasi, A.M.; El-Ansary, O.D.; Zin El-Abedin, K.T.; Yessoufou, K. Polyphenol Profile and Pharmaceutical Potential of Quercus spp. Bark Extracts. Plants 2019, 8, 486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elansary, H.O.; Salem, M.Z.M.; Ashmawy, N.A.; Yessoufou, K.; El-Settawy, A.A.A. In vitro antibacterial, antifungal and antioxidant activities of Eucalyptus spp. leaf extracts related to phenolic composition. Nat. Prod. Res. 2017, 31, 2927–2930. [Google Scholar] [CrossRef] [PubMed]
- Elansary, H.O.; Szopa, A.; Kubica, P.; Ekiert, H.; Ali, H.M.; Elshikh, M.S.; Abdel-Salam, E.M.; El-Esawi, M.; El-Ansary, D.O. Bioactivities of Traditional Medicinal Plants in Alexandria. Evid. Based Complement. Alternat. Med. 2018, 2018, 1463579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elansary, H.O.; Mahmoud, A.E.; El-Ansary, D.O.; Mattar, M.A. Effects of Water Stress and Modern Biostimulants on Growth and Quality Characteristics of Mint. Agronomy 2019, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Elansary, H.O.; Szopa , A.; Klimek-Szczykutowicz, M.; Jafernik, K.; Ekiert, H.; Mahmoud, E.A.; Barakat, A.A.; El-Ansary, D.O. Mammillaria Species—Polyphenols Studies and Anti-Cancer, Anti-Oxidant, and Anti-Bacterial Activities. Molecules 2019, 25, 131. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Morphological parameters and essential oil ratio in treated plants. A + E: A. nodosum + E. maxima. Different letters on each column indicate significant different at p ≤ 0.05.
Figure 1.
Morphological parameters and essential oil ratio in treated plants. A + E: A. nodosum + E. maxima. Different letters on each column indicate significant different at p ≤ 0.05.

Figure 2.
Gas exchange parameters measurements in plants subjected to seaweed treatments. Different letters on columns indicate significant differences among treatments at p ≥ 0.05. A + E: A. nodosum + E. maxima.
Figure 2.
Gas exchange parameters measurements in plants subjected to seaweed treatments. Different letters on columns indicate significant differences among treatments at p ≥ 0.05. A + E: A. nodosum + E. maxima.

Figure 3.
Showing the GC-MS of R. graveolens.

Figure 4.
Response of different cancer cells in the apoptotic cell population assay using flow cytometry to seaweed treatments, 2-undecanone, and 2-nonanone.
Figure 4.
Response of different cancer cells in the apoptotic cell population assay using flow cytometry to seaweed treatments, 2-undecanone, and 2-nonanone.

Figure 5.
Effect of seaweed treatments, 2-undecanone, and 2-nonanone (IC50) on the activity of caspase 3/7. The activity was expressed as a percentage (%) of untreated cells. Different letter on columns indicate significant differences within each treatment at p ≥ 0.05. A + E: A. nodosum + E. maxima.
Figure 5.
Effect of seaweed treatments, 2-undecanone, and 2-nonanone (IC50) on the activity of caspase 3/7. The activity was expressed as a percentage (%) of untreated cells. Different letter on columns indicate significant differences within each treatment at p ≥ 0.05. A + E: A. nodosum + E. maxima.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Essential oil main chemical constituents (means expressed in percentage) of R. graveolens as a response to seaweed extract treatments. Different letters indicate significant differences among treatments within the same row at p ≥ 0.05. A + E: A. nodosum + E. maxima.
Table 1.
Essential oil main chemical constituents (means expressed in percentage) of R. graveolens as a response to seaweed extract treatments. Different letters indicate significant differences among treatments within the same row at p ≥ 0.05. A + E: A. nodosum + E. maxima.
| Identified Compounds | Control | A. nodosum | E. maxima | A + E | |||
|---|---|---|---|---|---|---|---|
| RI | LRI | Standard Identification | |||||
| 1096 | 1096 | + | 2-nonanone | 17.01 ± 0.1 d | 19.11 ± 0.3 b | 18.21 ± 0.3 c | 18.09 ± 0.5 a |
| 1294 | 1294 | + | 2-undecanone | 49.66 ± 0.5 c | 54.38 ± 0.7 b | 53.1 ± 0.9 b | 61.9 ± 0.5 a |
Table 2.
In vitro antiproliferative activity, IC50 (µg mL−1), of R. graveolens EOs as affected by seaweed extracts (SWE) treatments.
Table 2.
In vitro antiproliferative activity, IC50 (µg mL−1), of R. graveolens EOs as affected by seaweed extracts (SWE) treatments.
| MCF-7 | HeLa | Jurkat | T24 | HT-29 | HEK-293 | |
|---|---|---|---|---|---|---|
| Control | 20.11 ± 0.9 | 6.5 ± 0.1 | 26.03 ± 0.1 | 95.3 ± 1.7 | 122.18 ± 2.3 | >200 |
| A. nodosum | 15.0 ± 0.3 | 5.24 ± 0.2 | 19.5 ± 0.8 | 83.85 ± 2.4 | 90.14 ± 3.8 | >200 |
| E. maxima | 16.7 ± 0.1 | 5.9 ± 0.3 | 20.46 ± 0.7 | 85.52 ± 3.1 | 101.17 ± 1.4 | >200 |
| A + E | 8.11 ± 0.7 | 4.3 ± 0.1 | 17.05 ± 0.3 | 82.2 ± 1.9 | 70.53 ± 2.1 | >200 |
| 2-undecanone | 4.21 ± 0.3 | 2.1 ± 0.1 | 8.32 ± 0.2 | 41.5 ± 0.7 | 33.26 ± 1.5 | >200 |
| 2-nonanone | 7.02 ± 0.2 | 3.2 ± 0.1 | 14.15 ± 0.1 | 64.1 ± 2.1 | 57.33 ± 2.2 | >200 |
| Vinblastine sulfate | ‒ | 2.4 ± 0.07 | 0.1 ± 0.03 | 61.44 ± 2.1 | 20.3 ± 0.8 | 50.3 ± 1.4 |
| Taxol | 0.08 ± 0.005 | ‒ | ‒ | ‒ | ‒ | ‒ |
Table 3.
Minimum inhibitory (MIC, mg mL−1) and bactericidal concentrations (MBC, mg mL−1) of R. graveolens EOs as affected by SWE treatments.
Table 3.
Minimum inhibitory (MIC, mg mL−1) and bactericidal concentrations (MBC, mg mL−1) of R. graveolens EOs as affected by SWE treatments.
| Bacteria | Control MIC MBC | A. nodosum MIC MBC | E. maxima MIC MBC | A + E MIC MBC | 2-Undecanone | 2-Nonanone | Streptomycin | Ampicillin |
|---|---|---|---|---|---|---|---|---|
| Bacillus cereus | 0.21 ± 0.01 | 0.13 ± 0.01 | 0.16 ± 0.01 | 0.07± 0.01 | 0.04± 0.01 | 0.06± 0.01 | 0.07 ± 0.02 | 0.09 ± 0.01 |
| 0.43 ± 0.03 | 0.29 ± 0.01 | 0.37± 0.01 | 0.16 ± 0.01 | 0.08 ± 0.01 | 0.13 ± 0.01 | 0.15 ± 0.02 | 0.18 ± 0.01 | |
| Dickeya solani | 0.42 ± 0.01 | 0.33± 0.01 | 0.38 ± 0.01 | 0.29 ± 0.01 | 0.16 ± 0.02 | 0.24 ± 0.02 | 0.22 ± 0.02 | 0.31 ± 0.01 |
| 0.93 ± 0.03 | 0.81 ± 0.03 | 0.85 ± 0.03 | 0.71 ± 0.02 | 0.32 ± 0.03 | 0.55 ± 0.01 | 0.43 ± 0.03 | 0.56 ± 0.01 | |
| Escherichia coli | 0.10 ± 0.01 | 0.80 ± 0.01 | 0.90 ± 0.02 | 0.70 ± 0.02 | 0.31 ± 0.03 | 0.54 ± 0.02 | 0.10 ± 0.01 | 0.20 ± 0.01 |
| 0.23 ± 0.01 | 0.20 ± 0.01 | 0.21 ± 0.03 | 0.16 ± 0.03 | 0.16 ± 0.01 | 0.12 ± 0.03 | 0.25 ± 0.01 | 0.40 ± 0.01 | |
| Listeria monocytogenes | 0.21 ± 0.01 | 0.19 ± 0.01 | 0.20± 0.01 | 0.17 ± 0.01 | 0.09 ± 0.01 | 0.14 ± 0.01 | 0.13 ± 0.01 | 0.16 ± 0.01 |
| 0.45 ± 0.03 | 0.39 ± 0.01 | 0.42 ± 0.01 | 0.31 ± 0.01 | 0.16 ± 0.03 | 0.24 ± 0.01 | 0.27 ± 0.01 | 0.30 ± 0.01 | |
| Micrococcus flavus | 0.21 ± 0.02 | 0.18 ± 0.01 | 0.19 ± 0.01 | 0.17 ± 0.01 | 0.09 ± 0.01 | 0.13 ± 0.01 | 0.11 ± 0.01 | 0.10 ± 0.01 |
| 0.47 ± 0.03 | 0.39 ± 0.01 | 0.43 ± 0.01 | 0.32 ± 0.01 | 0.16 ± 0.01 | 0.26 ± 0.01 | 0.20 ± 0.01 | 0.20± 0.01 | |
| Pectobacterium atrosepticum | 0.31 ± 0.01 | 0.23 ± 0.01 | 0.26 ± 0.02 | 0.20 ± 0.00 | 0.10 ± 0.01 | 0.16 ± 0.00 | 0.08 ± 0.01 | 0.26± 0.03 |
| 0.71 ± 0.03 | 0.45 ± 0.01 | 0.49 ± 0.01 | 0.40 ± 0.01 | 0.20 ± 0.01 | 0.31 ± 0.01 | 0.16 ± 0.01 | 0.52 ± 0.03 | |
| P. c. subsp. carotovorum | 0.11 ± 0.01 | 0.09 ± 0.01 | 0.10 ± 0.01 | 0.70 ± 0.01 | 0.33 ± 0.01 | 0.53 ± 0.01 | 0.09 ± 0.01 | 0.21± 0.01 |
| 0.25 ± 0.01 | 0.18 ± 0.00 | 0.23 ± 0.01 | 0.15 ± 0.01 | 0.07 ± 0.01 | 0.12 ± 0.01 | 0.20 ± 0.01 | 0.45 ± 0.03 | |
| Pseudomonas aeruginosa | 0.35 ± 0.01 | 0.28 ± 0.01 | 0.33± 0.01 | 0.25 ± 0.01 | 0.12 ± 0.01 | 0.21 ± 0.01 | 0.07 ± 0.02 | 0.11 ± 0.01 |
| 0.73 ± 0.01 | 0.54 ± 0.03 | 0.63 ± 0.01 | 0.53 ± 0.03 | 0.26 ± 0.01 | 0.42 ± 0.03 | 0.15 ± 0.01 | 0.24 ± 0.01 | |
| Staphylococcus aureus | 0.10 ± 0.00 | 0.08 ± 0.01 | 0.09 ± 0.01 | 0.06 ± 0.01 | 0.03 ± 0.00 | 0.05 ± 0.01 | 0.23 ± 0.02 | 0.10 ± 0.01 |
| 0.21 ± 0.01 | 0.19 ± 0.03 | 0.21 ± 0.01 | 0.13 ± 0.03 | 0.06 ± 0.01 | 0.10 ± 0.03 | 0.43 ± 0.01 | 0.21 ± 0.03 |
carotovorum and E. coli were the most sensitive bacteria. The antibacterial activity of EO from plants treated with A + E, were comparable to the antibacterial activity of antibiotics. Pure 2-undecanone and 2-nonanone showed lower values than all EOs.
Table 4.
Minimum inhibitory (MIC, mg mL−1) and fungicidal concentration (MFC, mg mL−1) of R. graveolens EOs as affected by SWE treatments.
Table 4.
Minimum inhibitory (MIC, mg mL−1) and fungicidal concentration (MFC, mg mL−1) of R. graveolens EOs as affected by SWE treatments.
| Fungi | Control MIC MFC | A. nodosum MIC MFC | E. maxima MIC MFC | A + E MIC MFC | 2-Undecanone | 2-Nonanone | FLZ MIC MFC | KTZ MIC MFC |
|---|---|---|---|---|---|---|---|---|
| Aspergillus flavus | 0.24 ± 0.01 | 0.19 ± 0.02 | 0.21 ± 0.01 | 0.17 ± 0.01 | 0.8 ± 0.01 | 0.14 ± 0.01 | 0.11 ± 0.01 | 0.20 ± 0.01 |
| 0.54 ± 0.01 | 0.47 ± 0.03 | 0.49 ± 0.01 | 0.43 ± 0.01 | 0.21 ± 0.01 | 0.37 ± 0.03 | 0.23 ± 0.03 | 0.42 ± 0.01 | |
| Aspergillus ochraceus | 0.22 ± 0.01 | 0.18 ± 0.01 | 0.20 ± 0.03 | 0.18 ± 0.01 | 0.09 ± 0.01 | 0.15 ± 0.01 | 0.19 ± 0.01 | 0.21 ± 0.01 |
| 0.51 ± 0.02 | 0.45 ± 0.01 | 0.49 ± 0.01 | 0.32 ± 0.01 | 0.15 ± 0.01 | 0.26 ± 0.01 | 0.33 ± 0.01 | 0.40 ± 0.01 | |
| Aspergillus niger | 0.21 ± 0.01 | 0.18 ± 0.03 | 0.19 ± 0.01 | 0.17 ± 0.01 | 0.08 ± 0.01 | 0.13 ± 0.01 | 0.14 ± 0.01 | 0.10 ± 0.01 |
| 0.48 ± 0.03 | 0.39 ± 0.01 | 0.42 ± 0.03 | 0.34 ± 0.03 | 0.16 ± 0.01 | 0.26 ± 0.02 | 0.29 ± 0.01 | 0.20 ± 0.01 | |
| Penicillium funiculosum | 0.31 ± 0.01 | 0.26 ± 0.02 | 0.29 ± 0.01 | 0.24 ± 0.01 | 0.22 ± 0.03 | 0.20 ± 0.01 | 0.13 ± 0.01 | 2.10 ± 0.07 |
| 0.63 ± 0.03 | 0.58 ± 0.03 | 0.59 ± 0.09 | 0.55 ± 0.07 | 0.26 ± 0.02 | 0.41 ± 0.03 | 0.25 ± 0.01 | 3.58 ± 0.05 | |
| Penicillium ochrochloron | 0.15 ± 0.01 | 0.12 ± 0.01 | 0.13 ± 0.01 | 0.12 ± 0.01 | 0.06 ± 0.01 | 0.10 ± 0.01 | 0.20 ± 0.01 | 0.17 ± 0.01 |
| 0.35 ± 0.01 | 0.25 ± 0.03 | 0.27 ± 0.03 | 0.22 ± 0.01 | 0.10 ± 0.01 | 0.17 ± 0.01 | 0.31 ± 0.01 | 0.31 ± 0.01 | |
| Candida albicans | 0.32 ± 0.13 | 0.28 ± 0.03 | 0.29 ± 0.11 | 0.26 ± 0.02 | 0.12 ± 0.01 | 0.21 ± 0.01 | 0.10 ± 0.01 | 0.20 ± 0.01 |
| 0.73 ± 0.11 | 0.62 ± 0.05 | 0.67 ± 0.14 | 0.61 ± 0.06 | 0.30 ± 0.01 | 0.52 ± 0.02 | 0.20 ± 0.01 | 0.41 ± 0.01 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mahmoud, E.A.; Elansary, H.O.; El-Ansary, D.O.; Al-Mana, F.A. Elevated Bioactivity of Ruta graveolens against Cancer Cells and Microbes Using Seaweeds. Processes 2020, 8, 75. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8010075
AMA Style
Mahmoud EA, Elansary HO, El-Ansary DO, Al-Mana FA. Elevated Bioactivity of Ruta graveolens against Cancer Cells and Microbes Using Seaweeds. Processes. 2020; 8(1):75. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8010075
Chicago/Turabian StyleMahmoud, Eman A., Hosam O. Elansary, Diaa O. El-Ansary, and Fahed A. Al-Mana. 2020. "Elevated Bioactivity of Ruta graveolens against Cancer Cells and Microbes Using Seaweeds" Processes 8, no. 1: 75. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8010075
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.