Model-Assisted Bird Monitoring Based on Remotely Sensed Ecosystem Functioning and Atlas Data

 ,
, 
 , and
, and
Abstract
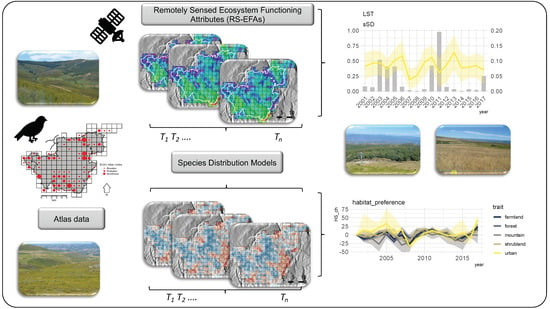
:
1. Introduction
2. Materials and Methods
2.1. Bird Data and Study Area
2.2. Satellite-Derived Products and EFAs
- (1)
- Enhanced Vegetation Index (EVI; MOD13Q1.006). EVI is an indicator of carbon gains since it is known to be more reliable in both low and high vegetation cover situations than Normalized Difference Vegetation Index (NDVI), and resistant to both soil influences, canopy background signals, and atmospheric effects on vegetation index values [33]. EVI values ranged from −1 to 1, with healthy vegetation generally holding values between 0.20 and 0.80.
- (2)
- Land-Surface Temperature (LST; MOD11A2.006). LST is a good indicator of the energy balance at the Earth’s surface, and one of the key parameters in the physics of land-surface processes from regional to global scales. In addition, LST is directly linked to the primary environmental regimes and to habitat suitability attributes (e.g., productivity, vegetation structure, land-cover type; [34]). Temperatures (LST) ranged from −25 °C to 45 °C.
- (3)
- Albedo (ALB; MCD43A3.006). ALB is surrogate for surface properties such as the extent and nature of the vegetation cover, and it is affected by the change of the land-surface bio-physical factors such as vegetation, LST and soil moisture [35]. ALB values ranged from 0 to 1 (fresh snow and bare soil usually fall around 0.9).
2.3. Species Distribution Models
2.4. Inter-Annual Predictions of Bird Communities
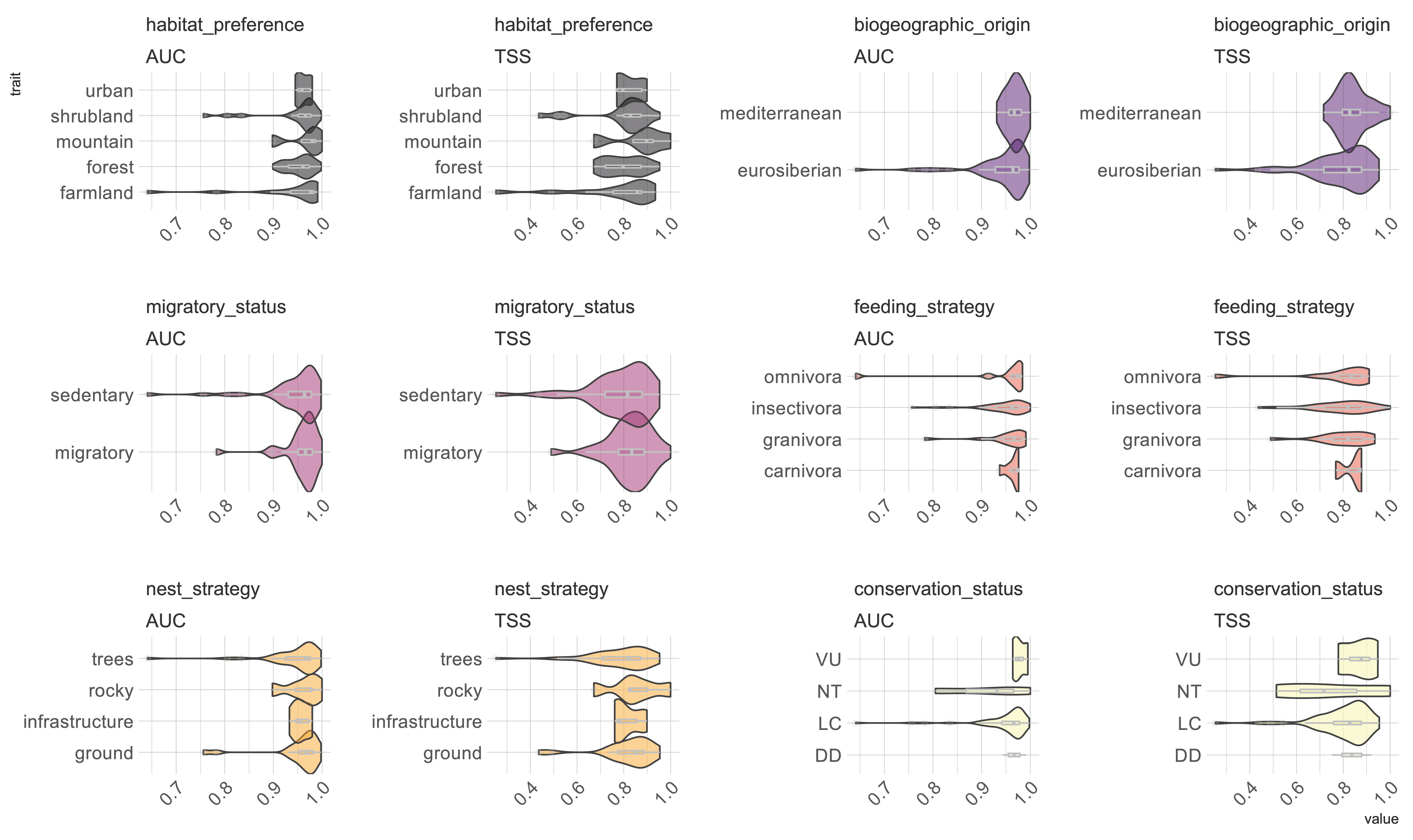
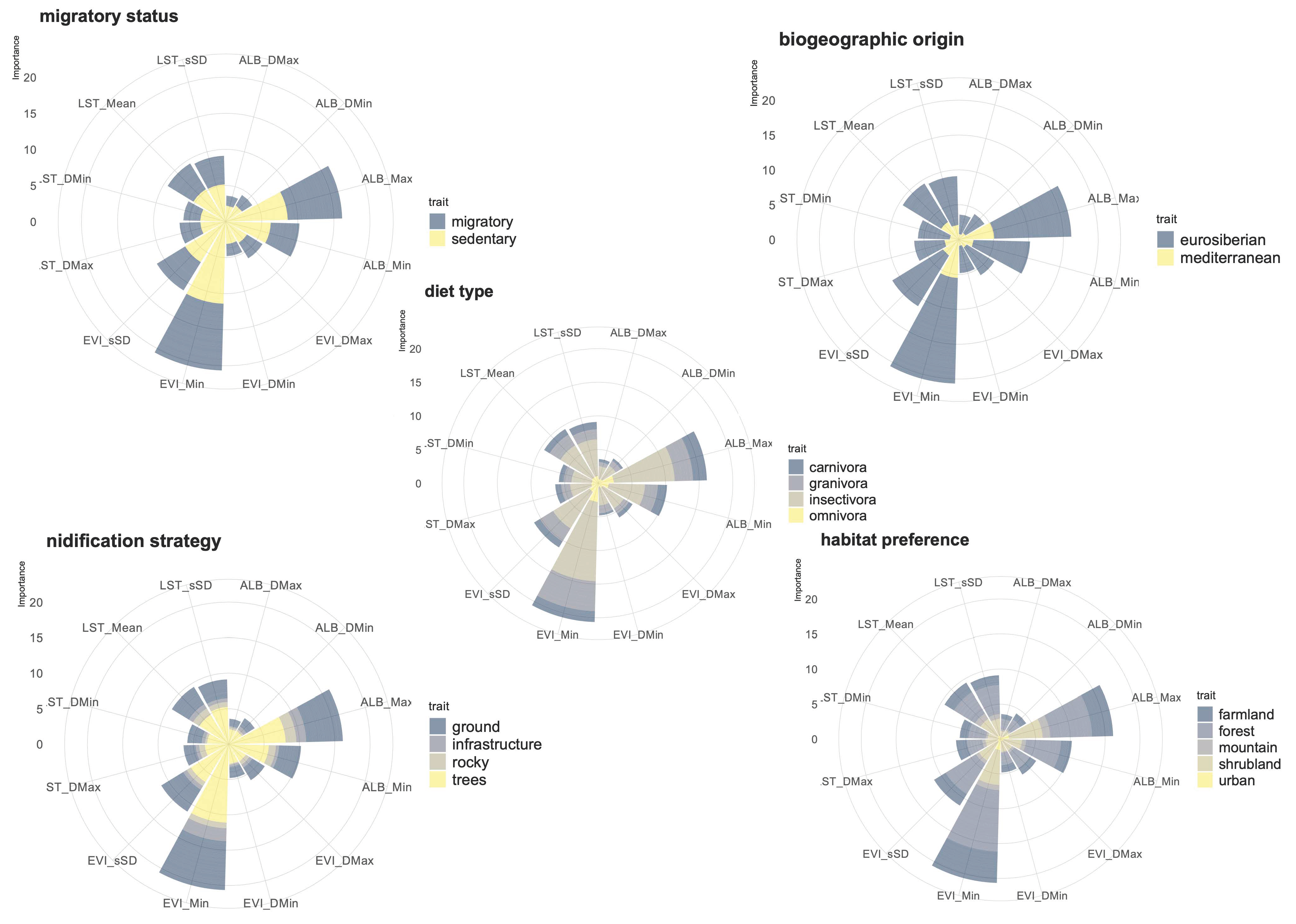
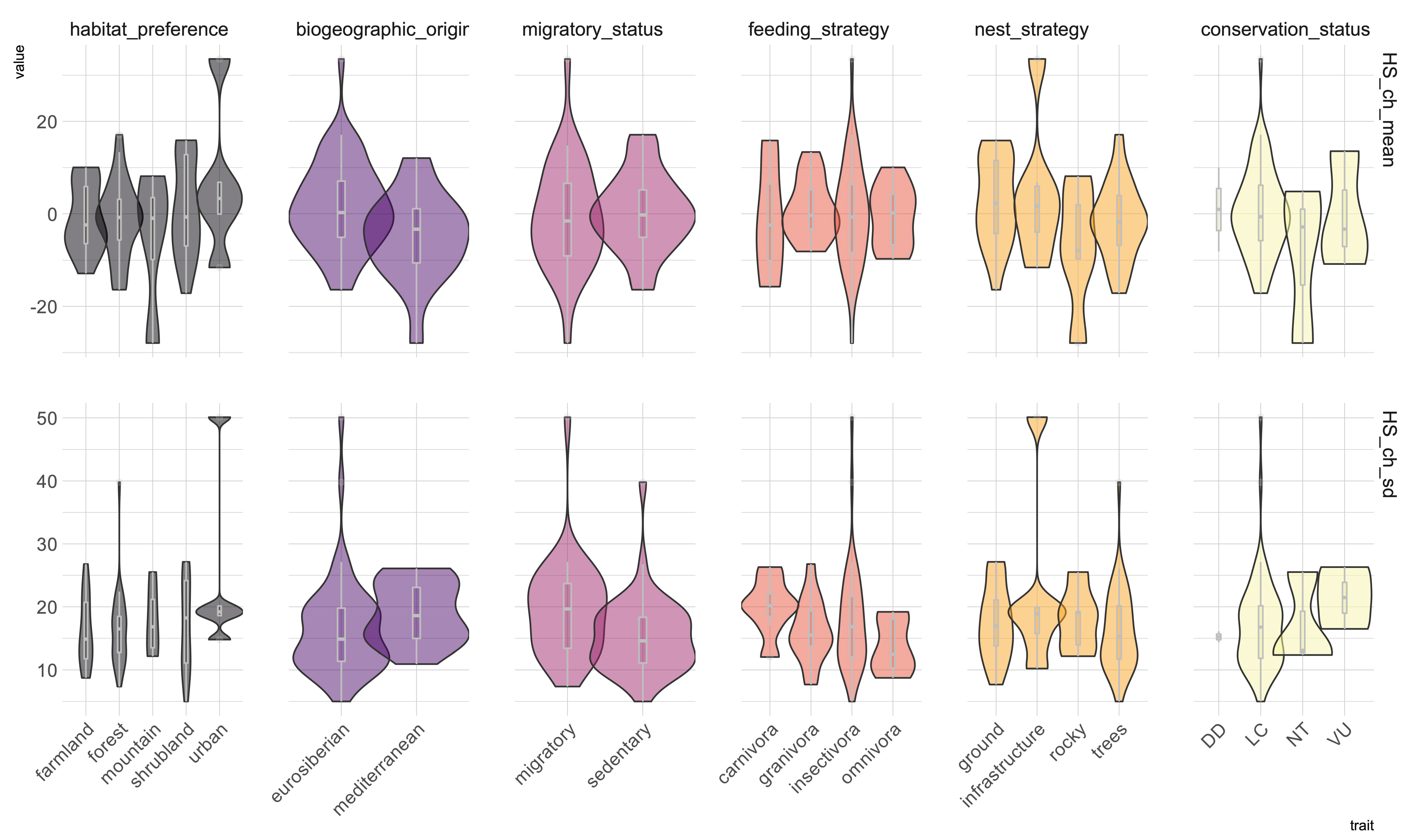
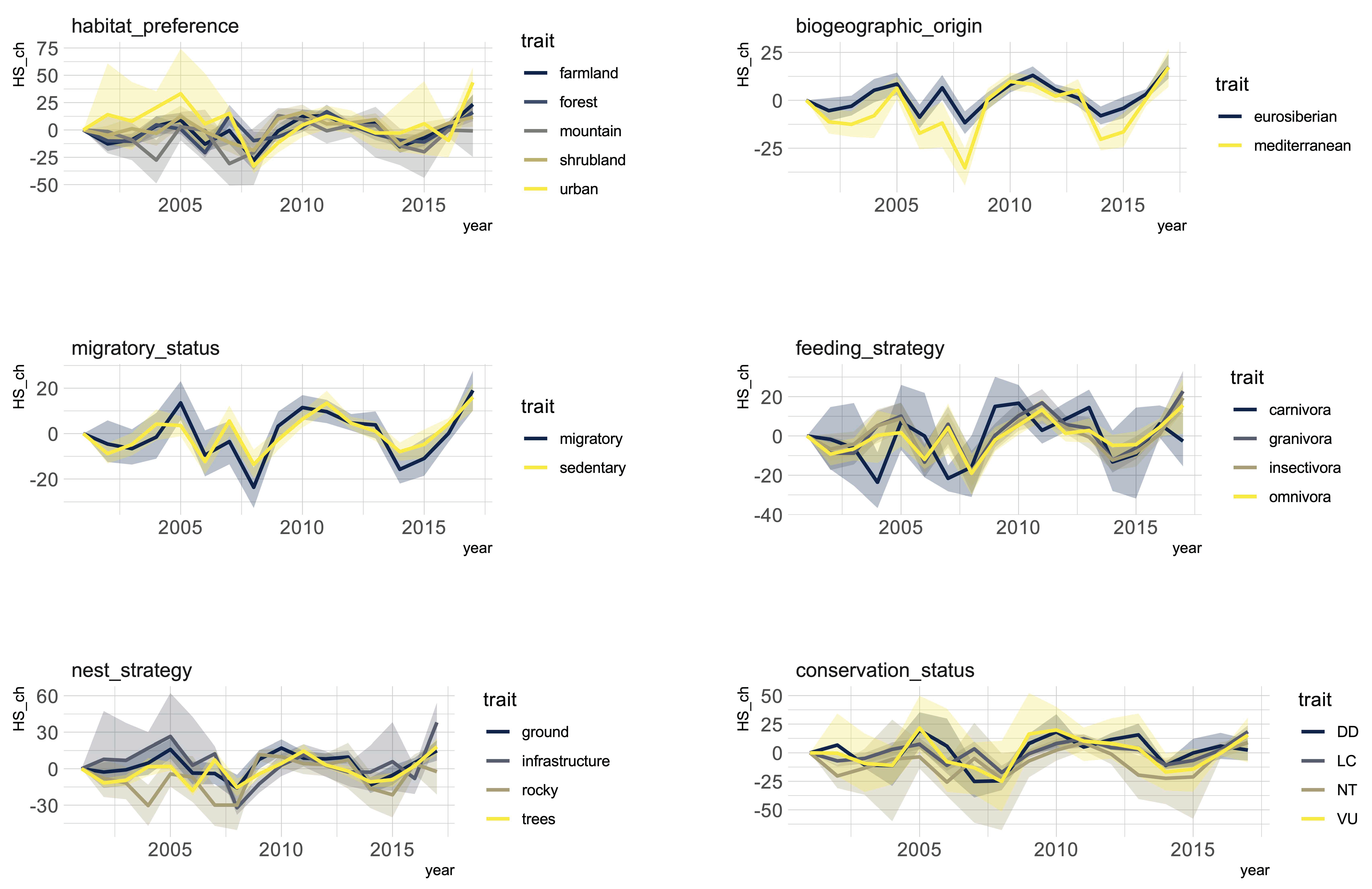
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Convention on Biological Diversity. CBD Report of the Tenth Meeting of the Conference of the Parties to the Convention on Biological Diversity; Technical Report; Convention on Biological Diversity: Nagoya, Japan, 2010. [Google Scholar]
- Pereira, H.M.; Ferrier, S.; Walters, M.; Geller, G.N.; Jongman, R.H.G.; Scholes, R.J.; Bruford, M.W.; Brummitt, N.; Butchart, S.H.M.; Cardoso, A.C.; et al. Essential Biodiversity Variables. Science 2013, 339, 277–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kissling, W.D.; Ahumada, J.A.; Bowser, A.; Fernandez, M.; Fernández, N.; García, E.A.; Guralnick, R.P.; Isaac, N.J.B.; Kelling, S.; Los, W.; et al. Building essential biodiversity variables (EBVs) of species distribution and abundance at a global scale. Biol. Rev. 2018, 93, 600–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jetz, W.; McGeoch, M.A.; Guralnick, R.; Ferrier, S.; Beck, J.; Costello, M.J.; Fernandez, M.; Geller, G.N.; Keil, P.; Merow, C.; et al. Essential biodiversity variables for mapping and monitoring species populations. Nat. Ecol. Evol. 2019, 3, 539–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Modell. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Honrado, P.; Pereira, H.M.; Guisan, A. Fostering integration between biodiversity monitoring and modelling. J. Appl. Ecol. 2016, 53, 1299–1304. [Google Scholar] [CrossRef]
- Schmeller, D.S.; Weatherdon, L.V.; Loyau, A.; Bondeau, A.; Brotons, L.; Brummitt, N.; Geijzendorffer, I.R.; Haase, P.; Kuemmerlen, M.; Martin, C.S.; et al. A suite of essential biodiversity variables for detecting critical biodiversity change. Biol. Rev. 2017, 93, 55–71. [Google Scholar] [CrossRef]
- O’Connor, B.; Secades, C.; Penner, J.; Sonnenschein, R.; Skidmore, A.; Burgess, N.D.; Hutton, J.M. Earth observation as a tool for tracking progress towards the Aichi Biodiversity Targets. Remote Sens. Ecol. Conserv. 2015, 1, 19–28. [Google Scholar] [CrossRef]
- He, K.S.; Bradley, B.A.; Cord, A.F.; Rocchini, D.; Tuanmu, M.-N.; Schmidtlein, S.; Turner, W.; Wegmann, M.; Pettorelli, N. Will remote sensing shape the next generation of species distribution models? Remote Sens. Ecol. Conserv. 2015, 1, 4–18. [Google Scholar] [CrossRef] [Green Version]
- Regos, A.; Gagne, L.; Alcaraz-Segura, D.; Honrado, J.P.; Domínguez, J. Effects of species traits and environmental predictors on performance and transferability of ecological niche models. Sci. Rep. 2019, 9, 4221. [Google Scholar] [CrossRef]
- Leitão, P.J.; Santos, M.J. Improving models of species ecological niches: A remote sensing overview. Front. Ecol. Evol. 2019, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Scherrer, D.; Guisan, A. Ecological indicator values reveal missing predictors of species distributions. Sci. Rep. 2019, 9, 3061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regos, A.; Vidal, M.; Lorenzo, M.; Domínguez, J. Integrating intraseasonal grassland dynamics in cross-scale distribution modeling to support waterbird recovery plans. Conserv. Biol. 2020, 34, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Pöyry, J.; Böttcher, K.; Fronzek, S.; Gobron, N.; Leinonen, R.; Metsämäki, S.; Virkkala, R. Predictive power of remote sensing versus temperature-derived variables in modelling phenology of herbivorous insects. Remote Sens. Ecol. Conserv. 2018, 4, 113–126. [Google Scholar] [CrossRef]
- Fernández, N.; Román, J.; Delibes, M. Variability in primary productivity determines metapopulation dynamics. Proc. R. Soc. B Biol. Sci. 2016, 283, 20152998. [Google Scholar] [CrossRef] [Green Version]
- Alcaraz, D.; Paruelo, J.; Cabello, J. Identification of current ecosystem functional types in the Iberian Peninsula. Glob. Ecol. Biogeogr. 2006, 15, 200–212. [Google Scholar] [CrossRef]
- Paruelo, J.M.; Burke, I.C.; Lauenroth, W.K. Land-use impact on ecosystem functioning in eastern Colorado, USA. Glob. Chang. Biol. 2001, 7, 631–639. [Google Scholar] [CrossRef]
- Alcaraz-Segura, D.; Lomba, A.; Sousa-Silva, R.; Nieto-Lugilde, D.; Alves, P.; Georges, D.; Vicente, J.R.; Honrado, J.P. Potential of satellite-derived ecosystem functional attributes to anticipate species range shifts. Int. J. Appl. Earth Obs. Geoinf. 2017, 57, 86–92. [Google Scholar] [CrossRef]
- Gonçalves, J.; Alves, P.; Pôças, I.; Marcos, B.; Sousa-Silva, R.; Lomba, Â.; Honrado, J.P. Exploring the spatiotemporal dynamics of habitat suitability to improve conservation management of a vulnerable plant species. Biodivers. Conserv. 2016, 25, 2867–2888. [Google Scholar] [CrossRef]
- Requena-Mullor, J.M.; López, E.; Castro, A.J.; Cabello, J.; Virgós, E.; González-Miras, E.; Castro, H. Modeling spatial distribution of European badger in arid landscapes: An ecosystem functioning approach. Landsc. Ecol. 2014, 29, 843–855. [Google Scholar] [CrossRef]
- Arenas-Castro, S.; Regos, A.; Gonçalves, J.F.; Alcaraz-Segura, D.; Honrado, J.P. Remotely Sensed Variables of Ecosystem Functioning Support Robust Predictions of Abundance Patterns for Rare Species. Remote Sens. 2019, 11, 2086. [Google Scholar] [CrossRef] [Green Version]
- Arenas-Castro, S.; Gonçalves, J.; Alves, P.; Alcaraz-Segura, D.; Honrado, J.P. Assessing the multi-scale predictive ability of ecosystem functional attributes for species distribution modelling. PLoS ONE 2018, 13, e0199292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syphard, A.D.; Franklin, J. Differences in spatial predictions among species distribution modeling methods vary with species traits and environmental predictors. Ecography 2009, 32, 907–918. [Google Scholar] [CrossRef]
- Wogan, G.O.U. Life history traits and niche instability impact accuracy and temporal transferability for historically calibrated distribution models of North American birds. PLoS ONE 2016, 11, e0151024. [Google Scholar] [CrossRef]
- Dobrowski, S.Z.; Thorne, J.H.; Greenberg, J.A.; Safford, H.D.; Mynsberge, A.R.; Crimmins, S.M.; Swanson, A.K. Modeling plant ranges over 75 years of climate change in California, USA: Temporal transferability and species traits. Ecol. Monogr. 2011, 81, 241–257. [Google Scholar] [CrossRef] [Green Version]
- Gruber, B.; Evans, D.; Henle, K.; Bauch, B.; Schmeller, D.; Dziock, F.; Henry, P.-Y.; Szabolcs, L.; Margules, C.; Dormann, C. “Mind the gap!”—How well does Natura 2000 cover species of European interest? Nat. Conserv. 2012, 3, 45–62. [Google Scholar] [CrossRef]
- Rodríguez-Lado, L.; Tapia, L.; Pérez, M.; Taboada, T.; Martínez-Capel, F.; Macías, F. Atlas Digital de Propiedades de Suelos de Galicia; University of Santiago de Compostela: Santiago, Spain, 2016. [Google Scholar]
- Tapia, L.; Regos, A.; Gil-Carrera, A.; Domínguez, J. Unravelling the response of diurnal raptors to land use change in a highly dynamic landscape in northwestern Spain: An approach based on satellite earth observation data. Eur. J. Wildl. Res. 2017, 63, 40. [Google Scholar] [CrossRef]
- Abel, D.; Regos, A.; Tapia, L.; Vidal, M.; Domínguez, J. Riqueza, abundancia e distribución de aves nidificantes nas Zonas de Especial Conservación (ZECs) Macizo Central e Bidueiral de Montederramo. Nov. Acta Cient. Compostel. 2019, 26, 109–123. [Google Scholar]
- Paruelo, J.M.; Jobbágy, E.G.; Sala, O.E. Current distribution of ecosystem functional types in temperate South America. Ecosystems 2001, 4, 683–698. [Google Scholar] [CrossRef]
- Cabello, J.; Fernández, N.; Alcaraz-Segura, D.; Oyonarte, C.; Piñeiro, G.; Altesor, A.; Delibes, M.; Paruelo, J.M. The ecosystem functioning dimension in conservation: Insights from remote sensing. Biodivers. Conserv. 2012, 21, 3287–3305. [Google Scholar] [CrossRef] [Green Version]
- Potter, C.; Gross, P.; Genovese, V.; Smith, M.L. Net primary productivity of forest stands in New Hampshire estimated from Landsat and MODIS satellite data. Carbon Balance Manag. 2007, 2, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cord, A.; Rödder, D. Inclusion of habitat availability in species distribution models through multi-temporal remote sensing data? Towards multifunctional agricultural landscapes in Europe: Assessing and governing synergies between food production, biodiversity, and ecosystem. Ecol. Appl. 2011, 21, 3285–3298. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, X.; Novillo, C.J.; Arrogante-funes, P.; Vázquez-jiménez, R.; Maestre, F.T. Albedo estimated from remote sensing correlates with ecosystem multifunctionality in global drylands. J. Arid Environ. 2018, 157, 116–123. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Van Proosdij, A.S.J.J.; Sosef, M.S.M.M.; Wieringa, J.J.; Raes, N. Minimum required number of specimen records to develop accurate species distribution models. Ecography 2016, 6, 542–552. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Townsend Peterson, A. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Stockwell, D.R.B.; Peterson, A.T. Effects of sample size on accuracy of species distribution models. Ecol. Modell. 2002, 148, 1–13. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD–A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Araújo, M.B.; New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- Tapia, L.; Regos, A.; Gil-Carrera, A.; Dominguez, J. Assessing the temporal transferability of raptor distribution models: Implications for conservation. Bird Conserv. Int. 2018, 28, 375–389. [Google Scholar] [CrossRef]
- Yates, K.L.; Bouchet, P.J.; Caley, M.J.; Mengersen, K.; Randin, C.F.; Parnell, S.; Fielding, A.H.; Bamford, A.J.; Ban, S.; Barbosa, A.M.; et al. Outstanding Challenges in the Transferability of Ecological Models. Trends Ecol. Evol. 2018, 33, 790–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenger, S.J.; Olden, J.D. Assessing transferability of ecological models: An underappreciated aspect of statistical validation. Methods Ecol. Evol. 2012, 3, 260–267. [Google Scholar] [CrossRef]
- Werkowska, W.; Márquez, A.L.; Real, R.; Acevedo, P. A practical overview of transferability in species distribution modeling. Environ. Rev. 2017, 25, 127–133. [Google Scholar] [CrossRef]
- Huang, Q.; Swatantran, A.; Dubayah, R.; Goetz, S.J. The influence of vegetation height heterogeneity on forest and woodland bird species richness across the United States. PLoS ONE 2014, 9, e103236. [Google Scholar] [CrossRef] [Green Version]
- Duro, D.C.; Girard, J.; King, D.J.; Fahrig, L.; Mitchell, S.; Lindsay, K.; Tischendorf, L. Predicting species diversity in agricultural environments using Landsat TM imagery. Remote Sens. Environ. 2014, 144, 214–225. [Google Scholar] [CrossRef]
- Morelli, F.; Pruscini, F.; Santolini, R.; Perna, P.; Benedetti, Y.; Sisti, D. Landscape heterogeneity metrics as indicators of bird diversity: Determining the optimal spatial scales in different landscapes. Ecol. Indic. 2013, 34, 372–379. [Google Scholar] [CrossRef]
- Thorup, K.; Tøttrup, A.P.; Willemoes, M.; Klaassen, R.H.G.; Strandberg, R.; Vega, M.L.; Dasari, H.P.; Araújo, M.B.; Wikelski, M.; Rahbek, C. Resource tracking within and across continents in long-distance bird migrants. Sci. Adv. 2017, 3, e1601360. [Google Scholar] [CrossRef] [Green Version]
- Shariatinajafabadi, M.; Wang, T.; Skidmore, A.K.; Toxopeus, A.G.; Kölzsch, A.; Nolet, B.A.; Exo, K.M.; Griffin, L.; Stahl, J.; Cabot, D. Migratory herbivorous waterfowl track satellite-derived green wave index. PLoS ONE 2014, 9, e108331. [Google Scholar] [CrossRef]
- Najafabadi, M.S.; Darvishzadeh, R.; Skidmore, A.K.; Kölzsch, A.; Vrieling, A.; Nolet, B.A.; Exo, K.M.; Meratnia, N.; Havinga, P.J.M.; Stahl, J.; et al. Satellite-versus temperature-derived green wave indices for predicting the timing of spring migration of avian herbivores. Ecol. Indic. 2015, 58, 322–331. [Google Scholar] [CrossRef] [Green Version]
- Shryock, B.; Marzluff, J.M.; Moskal, L.M. Urbanization alters the influence of weather and an index of forest productivity on avian community richness and guild abundance in the Seattle Metropolitan Area. Front. Ecol. Evol. 2017, 5, 40. [Google Scholar] [CrossRef] [Green Version]
- Suttidate, N.; Hobi, M.L.; Pidgeon, A.M.; Round, P.D.; Coops, N.C.; Helmers, D.P.; Keuler, N.S.; Dubinin, M.; Bateman, B.L.; Radeloff, V.C. Tropical bird species richness is strongly associated with patterns of primary productivity captured by the Dynamic Habitat Indices. Remote Sens. Environ. 2019, 232, 111306. [Google Scholar] [CrossRef]
- Toszogyova, A.; Storch, D. Global diversity patterns are modulated by temporal fluctuations in primary productivity. Glob. Ecol. Biogeogr. 2019, 28, 1827–1838. [Google Scholar] [CrossRef]
- Lehikoinen, A.; Brotons, L.; Calladine, J.; Campedelli, T.; Escandell, V.; Flousek, J.; Grueneberg, C.; Haas, F.; Harris, S.; Herrando, S.; et al. Declining population trends of European mountain birds. Glob. Chang. Biol. 2019, 25, 577–588. [Google Scholar] [CrossRef] [Green Version]
- Quintano, C.; Fernandez-Manso, A.; Marcos, E.; Calvo, L. Burn Severity and Post-Fire Land Surface Albedo Relationship in Mediterranean Forest Ecosystems. Remote Sens. 2019, 11, 2309. [Google Scholar] [CrossRef] [Green Version]
- Dintwe, K.; Okin, G.S.; Xue, Y. Fire-induced albedo change and surface radiative forcing in sub-Saharan Africa savanna ecosystems: Implications for the energy balance. J. Geophys. Res. 2017, 122, 6186–6201. [Google Scholar] [CrossRef] [Green Version]
- Rocha, A.V.; Shaver, G.R. Postfire energy exchange in arctic tundra: The importance and climatic implications of burn severity. Glob. Chang. Biol. 2011, 17, 2831–2841. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EFA Name | Acronyms | Description |
|---|---|---|
| Annual mean | Mean | Surrogate of annual total amount. |
| Maximum | Max | Annual maximum (indicator of the annual extremes). |
| Minimum | Min | Annual minimum (indicator of the annual extremes). |
| Annual coefficient of variation | sCV | A descriptor of the differences between seasons. |
| Annual standard deviation | sSD | A descriptor of the variations between seasons. |
| Date of maximum | DMx | Phenological indicator of the maximum growing season. |
| Date of minimum | DMi | Phenological indicator of the minimum growing season. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Regos, A.; Gómez-Rodríguez, P.; Arenas-Castro, S.; Tapia, L.; Vidal, M.; Domínguez, J. Model-Assisted Bird Monitoring Based on Remotely Sensed Ecosystem Functioning and Atlas Data. Remote Sens. 2020, 12, 2549. https://0-doi-org.brum.beds.ac.uk/10.3390/rs12162549
Regos A, Gómez-Rodríguez P, Arenas-Castro S, Tapia L, Vidal M, Domínguez J. Model-Assisted Bird Monitoring Based on Remotely Sensed Ecosystem Functioning and Atlas Data. Remote Sensing. 2020; 12(16):2549. https://0-doi-org.brum.beds.ac.uk/10.3390/rs12162549
Chicago/Turabian StyleRegos, Adrián, Pablo Gómez-Rodríguez, Salvador Arenas-Castro, Luis Tapia, María Vidal, and Jesús Domínguez. 2020. "Model-Assisted Bird Monitoring Based on Remotely Sensed Ecosystem Functioning and Atlas Data" Remote Sensing 12, no. 16: 2549. https://0-doi-org.brum.beds.ac.uk/10.3390/rs12162549