
May the Force Be with You (Or Not): The Immune System under Microgravity

 ,
,  , and
, and
Abstract
:1. Introduction
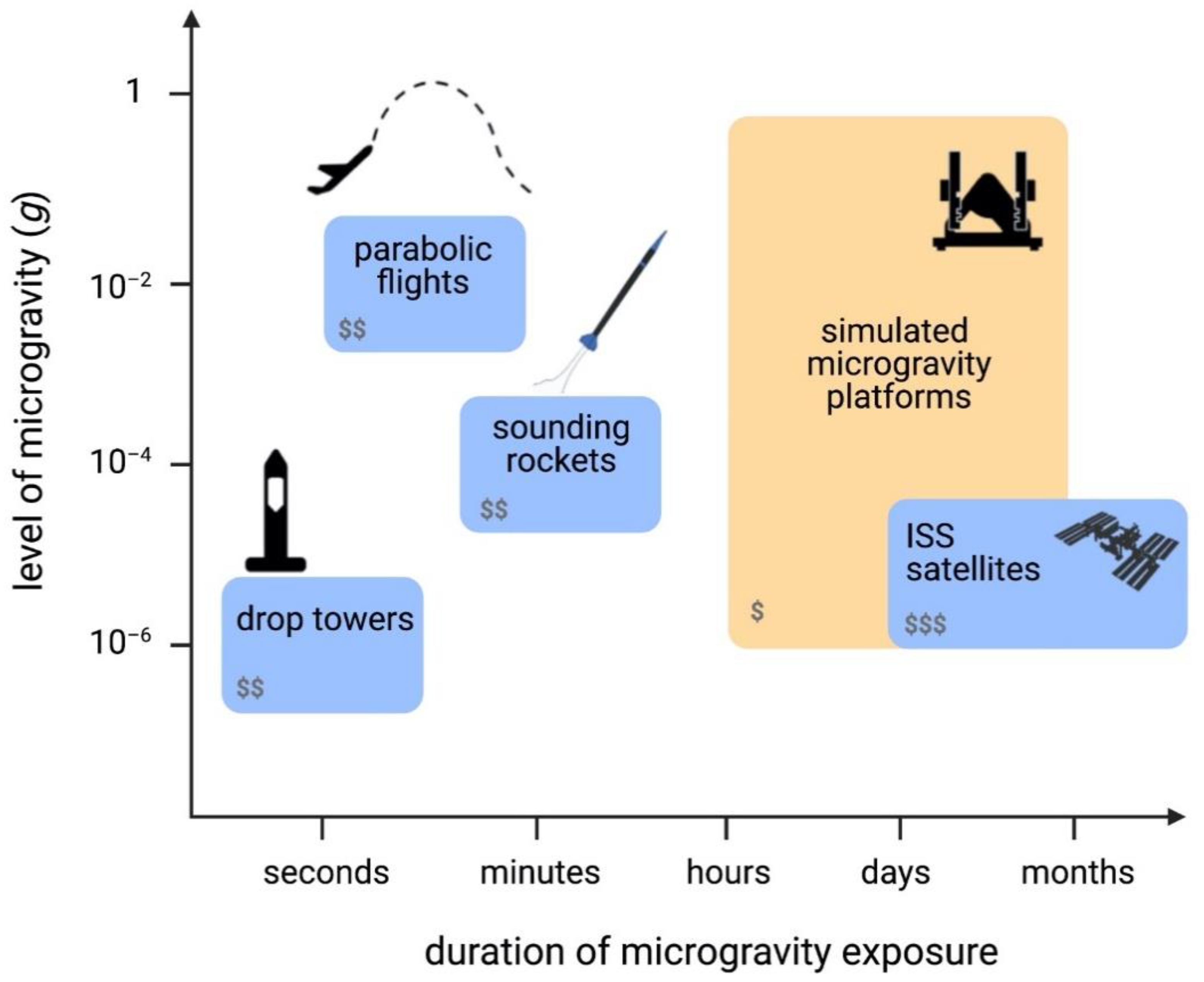
2. Real and Simulated Microgravity Platforms
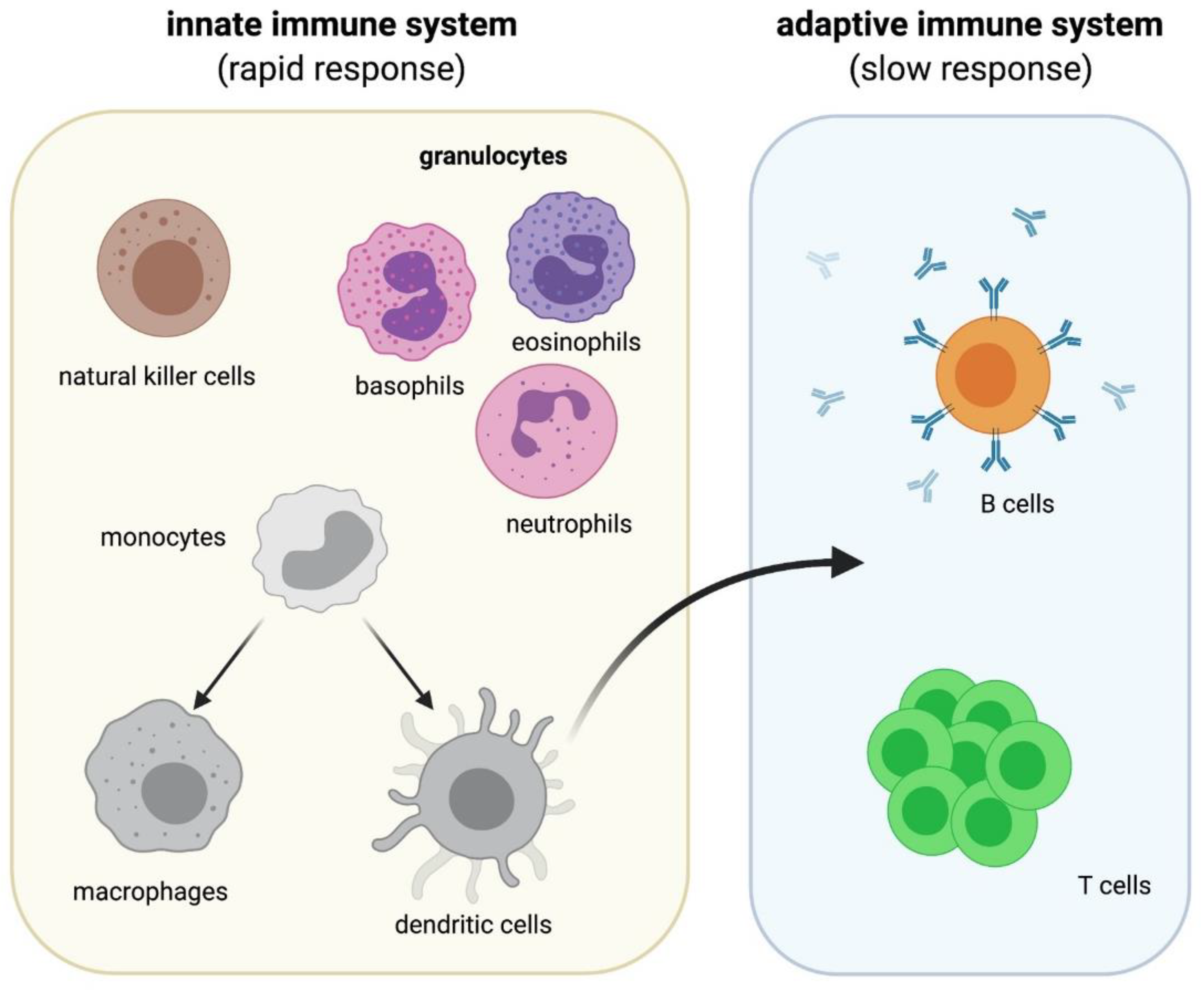
3. Immune Cells and Microgravity
3.1. Microgravity Studies on Cells of the Innate Immune System
3.1.1. Neutrophils
3.1.2. Natural Killer (NK) Cells
3.1.3. Monocytes
3.1.4. Macrophages
3.1.5. Dendritic Cells (DCs)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Type | Gravity Type | Platform | Cell Source | Observation | Ref. |
|---|---|---|---|---|---|
| Neutrophil | Real | Parabolic Flight | Human PBMC | Neutrophils are increased while all other immune cell subsets decrease | [44] |
| Spaceflight | Human PBMC | No change in neutrophil numbers before and after flight | [46] | ||
| Real/ Simulated | Spaceflight/ RWV | Human PBMC | Increase in neutrophils and neutrophil to lymphocyte ratio which could be used as a potential biomarker to monitor in flight health | [45] | |
| NK | Simulated | RWV | Human PBMC | Decrease in NK cell cytotoxicity related to a decrease in associated cytokine production and surface receptor expression | [47] |
| RWV | Human PBMC | NKG2D was found to be significantly reduced on the surface of NK cells and rescued with the addition of polysaccharides | [49] | ||
| Real | ISS | Human PBMC | Impaired NK cell function in terms of cytotoxic activity against leukemia K562 cell line | [48] | |
| Monocytes | Real | Spaceflight | Human PBMC | Total monocyte percentages were unchanged after a 13–16-day spaceflight | [50] |
| Parabolic Flight and Sounding Rocket | U937 | Microarray studies identified eight genes that remained stable during normal and microgravity conditions | [51] | ||
| Macrophages | Real | Spaceflight | J-111 | J-111 cells were found to have decreased cell motility and a reduction of F-actin | [55] |
| Spaceflight | U937 | PMA stimulated U937 cells had a disorganized cytoskeleton and decreased expression of CD18, CD36, and MHC-II | [54] | ||
| ISS | Human Primary M1 | M1 macrophages showed a decrease in ICAM-1 expression. No structural changes to actin or vimentin were seen and an increase in free fucose was also shown along with a decrease in cell surface bound fucose | [57] | ||
| Sounding Rocket | Human Primary M1 | Human primary M1 macrophages were found to have a significant reduction in cell volume, nuclear volume, and actin cytoskeleton as early as 4 s in microgravity | [56] | ||
| Real/ Simulated | Parabolic Flight/ 2D Clinostat | U937 | Microgravity conditions cause non-specific activation of U937 cells and PMA stimulated U937 cells are inhibited under these conditions | [26] | |
| Parabolic Flight/Spaceflight/2D Clinostat | U937/ Human Primary M2 | U937 human macrophages and human primary M2 macrophages had an increased expression of ICAM-1 | [54] | ||
| DCs | Simulated | RCCS | Human PBMC | pDC and mDC number are decreased under microgravity indicating that spaceflight could impair the development of these cells | [73] |
| RCCS | JAWS II DC | Prolonged culture of DCs in simulated microgravity reduced surface expression markers and the ability to activate T cells | [72] |
3.2. Microgravity Studies on Cells of the Adaptive Immune System
3.2.1. T Cells
3.2.2. B Cells
| Cell Type | Gravity Type | Platform | Cell Source | Observation | Ref. |
|---|---|---|---|---|---|
| T cells | Simulated | RWV | Human PBMC | Microgravity causes no impairment of DAG, or further downstream signaling, in CD4+ T cells | [85] |
| RCCS | Mouse Primary | CD25, CD69, IL-2 and IFNγ were all decreased in a time dependent manner from 24 to 72 h under simulated microgravity. CD4+ T cells were more susceptible to simulated microgravity effects of decreased proliferation than CD8+ T cells | [90] | ||
| RCCS | OT II mice | Long term culture of OT-II T cells results in resistance to activation by JAWS II DCs | [99] | ||
| RPM | Jurkat T cells | Decreased Ca2+ and ROS levels compared to 1 g controls | [101] | ||
| Real | Spaceflight | Human PBMC | Reduced expression of CD3, IL-2R and p44/42-MAPK-phosphorylation | [88] | |
| ISS | Human PBMC | Gene expression of miR-21 was suppressed in conA and anti-CD28 stimulated T cells | [92] | ||
| Spaceflight | OT II mice | 2-fold increase in OT-II cells in microgravity and an increase in IL-1b and IL-17 release when the cells were restimulated with OVA in vitro | [98] | ||
| Sounding Rocket/ Parabolic Flight | Jurkat T cells | Gene changes were found primarily in regulatory RNA | [93] | ||
| Parabolic Flight | Jurkat T cells | Levels of HIF1a remained relatively unchanged during microgravity exposure | [102] | ||
| Real/ Simulated | 2D Clinostat/ Parabolic Flight | Jurkat T cells | Increase in the phosphorylation of MAP kinases ERK-1/2, MEK, and p38 | [26] | |
| 2D Clinostat/ Parabolic Flight | Human PBMC/Jurkat T cells | Microgravity disrupted cell cycle regulatory proteins such as p21Waf1/Cip1, cdc2, and cdc25C | [86] | ||
| ISS/ RWV | Human PBMC | T cells had significantly downregulated gene expression of Rel/NF-B transcription factors | [87] | ||
| 2D/ parabolic | Human PBMC | Anti-CD28/conA activated T cells had decreased Zap-70 expression | [89] | ||
| Sounding Rocket/Parabolic Flight/2D Clinostat | Jurkat T cells | 5 genes remained unaltered in all microgravity conditions | [94] | ||
| Sounding Rocket/2D Clinostat | Jurkat T cells | 11 transcript clusters in non-activated Jurkat T cells were altered | [95] | ||
| Spaceflight/RWV and RPM | Mouse Primary | T cells had suppressed immune regulatory genes compared to controls | [96] | ||
| B cells | Simulated | RWV | HMy2.CIR | S decreased ion-radiation induced cell survival and increased apoptosis | [105] |
| Real | Parabolic Flight | Human PBMC | A decrease in B cells | [44] | |
| Spaceflight | Human PBMC | 15-day spaceflight showed no changes in B cell levels | [46] | ||
| Spaceflight | Mouse Primary | A significant reduction in B cells in the spleens of mice | [104] |
4. Future Directions and Discussion
4.1. Is There a Link between Microgravity and Mechanotransduction?
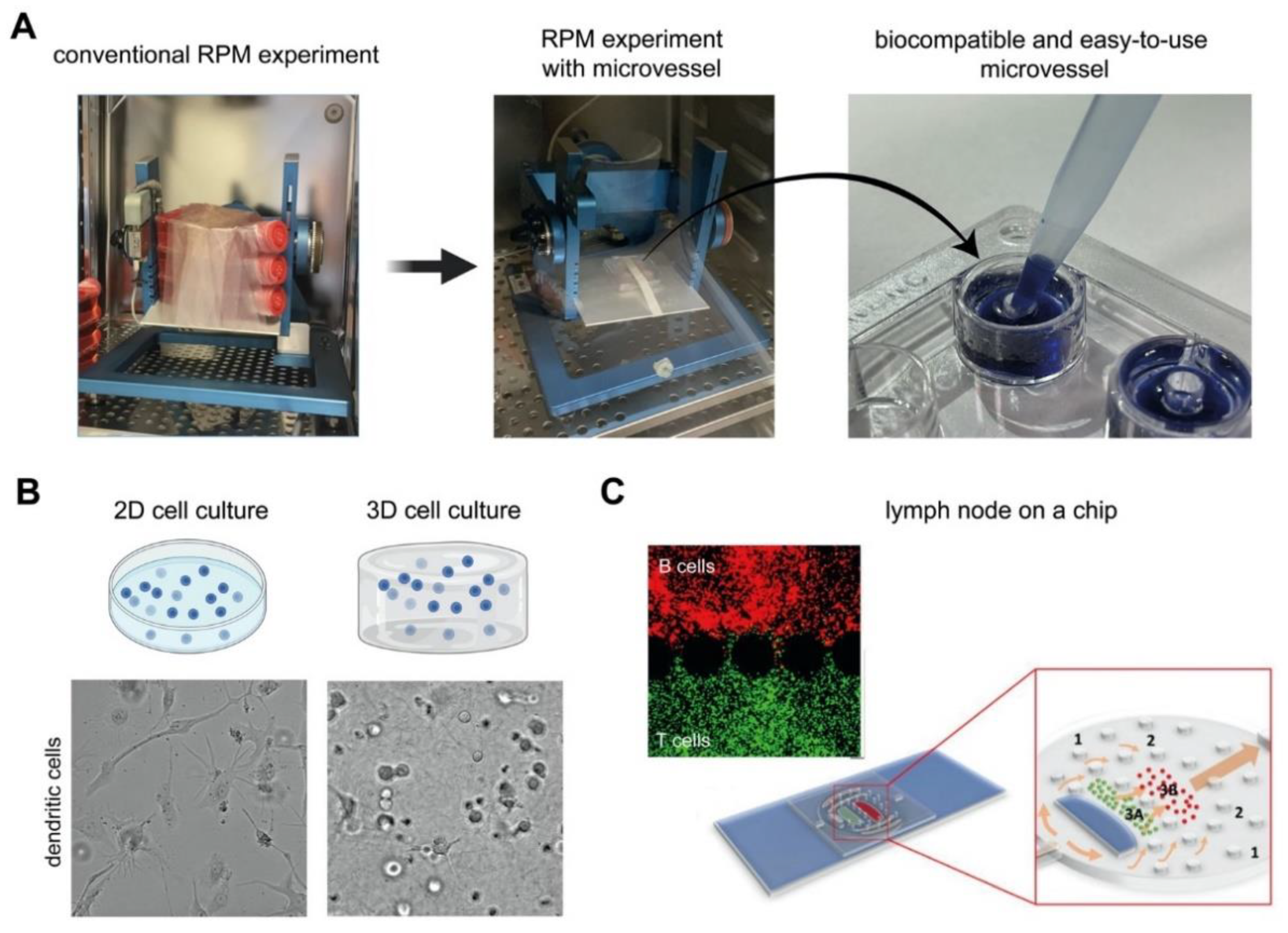
4.2. Technological Advances for Future Microgravity Research
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Ombergen, A.; Demertzi, A.; Tomilovskaya, E.; Jeurissen, B.; Sijbers, J.; Kozlovskaya, I.B.; Parizel, P.M.; Van de Heyning, P.H.; Sunaert, S.; Laureys, S.; et al. The effect of spaceflight and microgravity on the human brain. J. Neurol. 2017, 264, 18–22. [Google Scholar] [CrossRef] [Green Version]
- Grimm, D.; Grosse, J.; Wehland, M.; Mann, V.; Reseland, J.E.; Sundaresan, A.; Corydon, T.J. The impact of microgravity on bone in humans. Bone 2016, 87, 44–56. [Google Scholar] [CrossRef]
- Tanaka, K.; Nishimura, N.; Kawai, Y. Adaptation to microgravity, deconditioning, and countermeasures. J. Physiol. Sci. 2017, 67, 271–281. [Google Scholar] [CrossRef]
- White, R.J.; Averner, M. Humans in space. Nature 2001, 409, 1115–1118. [Google Scholar] [CrossRef] [PubMed]
- Sonnenfeld, G. The immune system in space and microgravity. Med. Sci. Sport. Exerc. 2002, 34, 2021–2027. [Google Scholar] [CrossRef] [PubMed]
- Prasad, B.; Grimm, D.; Strauch, S.M.; Erzinger, G.S.; Corydon, T.J.; Lebert, M.; Magnusson, N.E.; Infanger, M.; Richter, P.; Krüger, M. Influence of Microgravity on Apoptosis in Cells, Tissues, and Other Systems In Vivo and In Vitro. Int. J. Mol. Sci. 2020, 21, 9373. [Google Scholar] [CrossRef]
- Garrett-Bakelman, F.E.; Darshi, M.; Green, S.J.; Gur, R.C.; Lin, L.; Macias, B.R.; McKenna, M.J.; Meydan, C.; Mishra, T.; Nasrini, J.; et al. The NASA twins study: A multidimensional analysis of a year-long human spaceflight. Science 2019, 364. [Google Scholar] [CrossRef]
- Smith, J.K. Osteoclasts and microgravity. Life 2020, 10, 207. [Google Scholar] [CrossRef] [PubMed]
- Ikegame, M.; Hattori, A.; Tabata, M.J.; Kitamura, K.I.; Tabuchi, Y.; Furusawa, Y.; Maruyama, Y.; Yamamoto, T.; Sekiguchi, T.; Matsuoka, R.; et al. Melatonin is a potential drug for the prevention of bone loss during space flight. J. Pineal Res. 2019, 67, e12594. [Google Scholar] [CrossRef]
- Wu, C.H.; Ou, C.H.; Yen, I.C.; Lee, S.Y. 4-Acetylantroquinonol B Inhibits Osteoclastogenesis By Inhibiting the Autophagy Pathway in a Simulated Microgravity Model. Int. J. Mol. Sci. 2020, 21, 6971. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Lehar, A.; Meir, J.U.; Koch, C.; Morgan, A.; Warren, L.E.; Rydzik, R.; Youngstrom, D.W.; Chandok, H.; George, J.; et al. Targeting myostatin/activin A protects against skeletal muscle and bone loss during spaceflight. Proc. Natl. Acad. Sci. USA 2020, 117, 23942–23951. [Google Scholar] [CrossRef]
- Sotnezova, E.V.; Markina, E.A.; Andreeva, E.R.; Buravkova, L.B. Myeloid Precursors in the Bone Marrow of Mice after a 30-Day Space Mission on a Bion-M1 Biosatellite. Bull. Exp. Biol. Med. 2017, 162, 496–500. [Google Scholar] [CrossRef]
- Donaubauer, A.J.; Deloch, L.; Becker, I.; Fietkau, R.; Frey, B.; Gaipl, U.S. The influence of radiation on bone and bone cells—differential effects on osteoclasts and osteoblasts. Int. J. Mol. Sci. 2020, 21, 6377. [Google Scholar] [CrossRef] [PubMed]
- Zayzafoon, M.; Meyers, V.E.; McDonald, J.M. Microgravity: The immune response and bone. Immunol. Rev. 2005, 208, 267–280. [Google Scholar] [CrossRef] [PubMed]
- Patel, Z.S.; Brunstetter, T.J.; Tarver, W.J.; Whitmire, A.M.; Zwart, S.R.; Smith, S.M.; Huff, J.L. Red risks for a journey to the red planet: The highest priority human health risks for a mission to Mars. NPJ Microgravity 2020, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Prasanth, D.; Suresh, S.; Prathivadhi-Bhayankaram, S.; Mimlitz, M.; Zetocha, N.; Lee, B.; Ekpenyong, A. Microgravity modulates effects of chemotherapeutic drugs on cancer cell migration. bioRxiv 2019, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ahn, C.B.; Lee, J.H.; Han, D.G.; Kang, H.W.; Lee, S.H.; Lee, J.I.; Son, K.H.; Lee, J.W. Simulated microgravity with floating environment promotes migration of non-small cell lung cancers. Sci. Rep. 2019, 9, 14553. [Google Scholar] [CrossRef] [PubMed]
- Marshall-Goebel, K.; Damani, R.; Bershad, E.M. Brain Physiological Response and Adaptation during Spaceflight. Clin. Neurosurg. 2019, 85, E815–E821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overbey, E.G.; Paul, A.M.; da Silveira, W.A.; Tahimic, C.G.T.; Reinsch, S.S.; Szewczyk, N.; Stanbouly, S.; Wang, C.; Galazka, J.M.; Mao, X.W. Mice exposed to combined chronic low-dose irradiation and modeled microgravity develop long-term neurological sequelae. Int. J. Mol. Sci. 2019, 20, 4094. [Google Scholar] [CrossRef] [Green Version]
- Khossravi, E.A.; Hargens, A.R. Visual disturbances during prolonged space missions. Curr. Opin. Ophthalmol. 2021, 32, 69–73. [Google Scholar] [CrossRef]
- Crucian, B.; Simpson, R.J.; Mehta, S.; Stowe, R.; Chouker, A.; Hwang, S.A.; Actor, J.K.; Salam, A.P.; Pierson, D.; Sams, C. Terrestrial stress analogs for spaceflight associated immune system dysregulation. Brain. Behav. Immun. 2014, 39, 23–32. [Google Scholar] [CrossRef]
- Bradbury, P.; Wu, H.; Choi, J.U.; Rowan, A.E.; Zhang, H.; Poole, K.; Lauko, J.; Chou, J. Modeling the Impact of Microgravity at the Cellular Level: Implications for Human Disease. Front. Cell Dev. Biol. 2020, 8, 96. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Zhang, K.; Wei, D.; Tian, Y.; Gao, Y.; Chen, Z.; Qian, A. The impact of spaceflight and simulated microgravity on cell adhesion. Int. J. Mol. Sci. 2020, 21, 3031. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, T.; Horie, K.; Hinoi, E.; Hiraiwa, M.; Kato, A.; Maekawa, Y.; Takahashi, A.; Furukawa, S. How does spaceflight affect the acquired immune system? Jpn. Microgravity 2020, 6, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.K. IL-6 and the dysregulation of immune, bone, muscle, and metabolic homeostasis during spaceflight. npj Microgravity 2018, 4, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulsen, K.; Thiel, C.; Timm, J.; Schmidt, P.M.; Huber, K.; Tauber, S.; Hemmersbach, R.; Seibt, D.; Kroll, H.; Grote, K.-H.; et al. Microgravity-induced alterations in signal transduction in cells of the immune system. Acta Astronaut. 2010, 67, 1116–1125. [Google Scholar] [CrossRef]
- Choukèr, A.; Ullrich, O. The Immune System in Space: Are We Prepared? SpringerBriefs in Space Life Sciences; Springer International Publishing: Cham, Switzerland, 2016; ISBN 978-3-319-41464-5. [Google Scholar]
- Shetty, A.K. Introduction. Aging Cell 2004, 3, 331. [Google Scholar] [CrossRef]
- Secrier, M.; Schneider, R. Visualizing time-related data in biology, a review. Brief. Bioinform. 2014, 15, 771–782. [Google Scholar] [CrossRef] [Green Version]
- Ferranti, F.; Del Bianco, M.; Pacelli, C. Advantages and limitations of current microgravity platforms for space biology research. Appl. Sci. 2021, 11, 68. [Google Scholar] [CrossRef]
- Maier, J.A.M.; Cialdai, F.; Monici, M.; Morbidelli, L. The Impact of Microgravity and Hypergravity on Endothelial Cells. Biomed Res. Int. 2015, 2015, 1–13. [Google Scholar] [CrossRef]
- Wuest, S.L.; Stern, P.; Casartelli, E.; Egli, M. Fluid Dynamics Appearing during Simulated Microgravity Using Random Positioning Machines. PLoS ONE 2017, 12, e0170826. [Google Scholar] [CrossRef] [PubMed]
- Herranz, R.; Anken, R.; Boonstra, J.; Braun, M.; Christianen, P.C.M.; de Geest, M.; Hauslage, J.; Hilbig, R.; Hill, R.J.A.; Lebert, M.; et al. Ground-Based Facilities for Simulation of Microgravity: Organism-Specific Recommendations for Their Use, and Recommended Terminology. Astrobiology 2013, 13, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammond, T.G.; Hammond, J.M. Optimized suspension culture: The rotating-wall vessel. Am. J. Physiol. Physiol. 2001, 281, F12–F25. [Google Scholar] [CrossRef] [PubMed]
- Cyster, J.G.; Allen, C.D.C. B Cell Responses: Cell Interaction Dynamics and Decisions. Cell 2019, 177, 524–540. [Google Scholar] [CrossRef] [Green Version]
- Davenport, M.P.; Smith, N.L.; Rudd, B.D. Building a T cell compartment: How immune cell development shapes function. Nat. Rev. Immunol. 2020, 20, 499–506. [Google Scholar] [CrossRef]
- Chaplin, D.D. Overview of the immune response. J. Allergy Clin. Immunol. 2010, 125, S345. [Google Scholar] [CrossRef]
- Witzel, I.I.; Nasser, R.; Garcia-Sabaté, A.; Sapudom, J.; Ma, C.; Chen, W.; Teo, J.C.M. Deconstructing Immune Microenvironments of Lymphoid Tissues for Reverse Engineering. Adv. Healthc. Mater. 2019, 8, 1801126. [Google Scholar] [CrossRef]
- Tavassoli, M. Medical problems of space flight. Am. J. Med. 1986, 81, 850–854. [Google Scholar] [CrossRef]
- Shaw, A.C.; Goldstein, D.R.; Montgomery, R.R. Age-dependent dysregulation of innate immunity. Nat. Rev. Immunol. 2013, 13, 875–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frizinsky, S.; Haj-Yahia, S.; MacHnes Maayan, D.; Lifshitz, Y.; Maoz-Segal, R.; Offengenden, I.; Kidon, M.; Agmon-Levin, N. The innate immune perspective of autoimmune and autoinflammatory conditions. Rheumatology 2019, 58, VI1–VI8. [Google Scholar] [CrossRef] [Green Version]
- Kaur, I.; Simons, E.R.; Castro, V.A.; Mark Ott, C.; Pierson, D.L. Changes in neutrophil functions in astronauts. Brain. Behav. Immun. 2004, 18, 443–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichiki, A.T.; Gibson, L.A.; Jago, T.L.; Strickland, K.M.; Johnson, D.L.; Lange, R.D.; Allebban, Z. Effects of spaceflight on rat peripheral blood leukocytes and bone marrow progenitor cells. J. Leukoc. Biol. 1996, 60, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Stervbo, U.; Roch, T.; Kornprobst, T.; Sawitzki, B.; Grütz, G.; Wilhelm, A.; Lacombe, F.; Allou, K.; Kaymer, M.; Pacheco, A.; et al. Gravitational stress during parabolic flights reduces the number of circulating innate and adaptive leukocyte subsets in human blood. PLoS ONE 2019, 13, e020627. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.M.; Mhatre, S.D.; Cekanaviciute, E.; Schreurs, A.S.; Tahimic, C.G.T.; Globus, R.K.; Anand, S.; Crucian, B.E.; Bhattacharya, S. Neutrophil-to-Lymphocyte Ratio: A Biomarker to Monitor the Immune Status of Astronauts. Front. Immunol. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Cao, D.; Song, J.; Ling, S.; Niu, S.; Lu, L.; Cui, Z.; Li, Y.; Hao, S.; Zhong, G.; Qi, Z.; et al. Hematopoietic stem cells and lineage cells undergo dynamic alterations under microgravity and recovery conditions. FASEB J. 2019, 33, 6904–6918. [Google Scholar] [CrossRef]
- Li, Q.; Mei, Q.; Huyan, T.; Xie, L.; Che, S.; Yang, H.; Zhang, M.; Huang, Q. Effects of simulated microgravity on primary human NK cells. Astrobiology 2013, 13, 703–714. [Google Scholar] [CrossRef]
- Bigley, A.B.; Agha, N.H.; Baker, F.L.; Spielmann, G.; Kunz, H.E.; Mylabathula, P.L.; Rooney, B.V.; Laughlin, M.S.; Mehta, S.K.; Pierson, D.L.; et al. NK cell function is impaired during long-duration spaceflight. J. Appl. Physiol. 2019, 126, 842–853. [Google Scholar] [CrossRef]
- Huyan, T.; Li, Q.; Yang, H.; Jin, M.L.; Zhang, M.J.; Ye, L.J.; Li, J.; Huang, Q.S.; Yin, D.C. Protective effect of polysaccharides on simulated microgravity-induced functional inhibition of human NK cells. Carbohydr. Polym. 2014, 101, 819–827. [Google Scholar] [CrossRef]
- Crucian, B.; Stowe, R.; Quiriarte, H.; Pierson, D.; Sams, C. Monocyte phenotype and cytokine production profiles are dysregulated by short-duration spaceflight. Aviat. Sp. Environ. Med. 2011, 82, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Thiel, C.S.; Hauschild, S.; Tauber, S.; Paulsen, K.; Raig, C.; Raem, A.; Biskup, J.; Gutewort, A.; Hürlimann, E.; Unverdorben, F.; et al. Identification of reference genes in human myelomonocytic cells for gene expression studies in altered gravity. Biomed Res. Int. 2015, 2015, 363575. [Google Scholar] [CrossRef]
- Sapudom, J.; Mohamed, W.K.E.; Garcia-Sabaté, A.; Alatoom, A.; Karaman, S.; Mahtani, N.; Teo, J.C.M. Collagen Fibril Density Modulates Macrophage Activation and Cellular Functions during Tissue Repair. Bioengineering 2020, 7, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, D.Y.S.; Glim, J.E.; Stavenuiter, A.W.D.; Breur, M.; Heijnen, P.; Amor, S.; Dijkstra, C.D.; Beelen, R.H.J. Human macrophage polarization in vitro: Maturation and activation methods compared. Immunobiology 2014, 219, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, K.; Tauber, S.; Goelz, N.; Simmet, D.M.; Engeli, S.; Birlem, M.; Dumrese, C.; Karer, A.; Hunziker, S.; Biskup, J.; et al. Severe disruption of the cytoskeleton and immunologically relevant surface molecules in a human macrophageal cell line in microgravity—Results of an in vitro experiment on board of the Shenzhou-8 space mission. Acta Astronaut. 2014, 94, 277–292. [Google Scholar] [CrossRef] [Green Version]
- Meloni, M.A.; Galleri, G.; Pani, G.; Saba, A.; Pippia, P.; Cogoli-Greuter, M. Space flight affects motility and cytoskeletal structures in human monocyte cell line J-111. Cytoskeleton 2011, 68, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, K.; Tauber, S.; Dumrese, C.; Bradacs, G.; Simmet, D.M.; Gölz, N.; Hauschild, S.; Raig, C.; Engeli, S.; Gutewort, A.; et al. Regulation of ICAM-1 in cells of the monocyte/macrophage system in microgravity. Biomed Res. Int. 2015, 2015, 1–18. [Google Scholar] [CrossRef]
- Tauber, S.; Lauber, B.A.; Paulsen, K.; Layer, L.E.; Lehmann, M.; Hauschild, S.; Shepherd, N.R.; Polzer, J.; Segerer, J.; Thiel, C.S.; et al. Cytoskeletal stability and metabolic alterations in primary human macrophages in long-term microgravity. PLoS ONE 2017, 12, e0175599. [Google Scholar] [CrossRef] [Green Version]
- Thiel, C.S.; Tauber, S.; Lauber, B.; Polzer, J.; Seebacher, C.; Uhl, R.; Neelam, S.; Zhang, Y.; Levine, H.; Ullrich, O. Rapid Morphological and Cytoskeletal Response to Microgravity in Human Primary Macrophages. Int. J. Mol. Sci. 2019, 20, 2402. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Luo, H.; Zhu, L.; Yang, F.; Chu, Z.; Tian, H.; Feng, M.; Zhao, Y.; Shang, P. Microgravity inhibition of lipopolysaccharide-induced tumor necrosis factor-α expression in macrophage cells. Inflamm. Res. 2014, 63, 91–98. [Google Scholar] [CrossRef]
- Wang, C.; Chen, H.; Luo, H.; Zhu, L.; Zhao, Y.; Tian, H.; Wang, R.; Shang, P.; Zhao, Y. Microgravity activates p38 MAPK-C/EBPβ pathway to regulate the expression of arginase and inflammatory cytokines in macrophages. Inflamm. Res. 2015, 64, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Brungs, S.; Kolanus, W.; Hemmersbach, R. Syk phosphorylation—A gravisensitive step in macrophage signalling. Cell Commun. Signal. 2015, 13, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crucian, B.; Stowe, R.P.; Mehta, S.; Quiriarte, H.; Pierson, D.; Sams, C. Alterations in adaptive immunity persist during long-duration spaceflight. Jpn. Microgravity 2015, 1, 15013. [Google Scholar] [CrossRef] [PubMed]
- Thiel, C.S.; De Zélicourt, D.; Tauber, S.; Adrian, A.; Franz, M.; Simmet, D.M.; Schoppmann, K.; Hauschild, S.; Krammer, S.; Christen, M.; et al. Rapid adaptation to microgravity in mammalian macrophage cells. Sci. Rep. 2017, 7, 2266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanley, A.; Thompson, K.; Hynes, A.; Brakebusch, C.; Quondamatteo, F. NADPH Oxidase Complex-Derived Reactive Oxygen Species, the Actin Cytoskeleton, and Rho GTPases in Cell Migration. Antioxid. Redox Signal. 2014, 20, 2026–2042. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Tian, H.; Wang, P.; Li, L.; Zhang, Z.; Zhang, J.; Zhao, Y. Spaceflight and simulated microgravity suppresses macrophage development via altered RAS/ERK/NFκB and metabolic pathways. Cell. Mol. Immunol. 2020, 18, 1489–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Han, C.; Lu, Z.; Ge, P.; Cui, Y.; Zhao, D.; Yang, X.; Wu, B.; Qiang, L.; Zhang, Y.; et al. Simulated microgravity suppresses MAPK pathway-mediated innate immune response to bacterial infection and induces gut microbiota dysbiosis. FASEB J. 2020, 34, 14631–14644. [Google Scholar] [CrossRef] [PubMed]
- Chapuis, F.; Rosenzwajg, M.; Yagello, M.; Ekman, M.; Biberfeld, P.; Gluckman, J.C. Differentiation of human dendritic cells from monocytes in vitro. Eur. J. Immunol. 1997, 27, 431–441. [Google Scholar] [CrossRef]
- Tang-Huau, T.L.; Gueguen, P.; Goudot, C.; Durand, M.; Bohec, M.; Baulande, S.; Pasquier, B.; Amigorena, S.; Segura, E. Human in vivo-generated monocyte-derived dendritic cells and macrophages cross-present antigens through a vacuolar pathway. Nat. Commun. 2018, 9, 2570. [Google Scholar] [CrossRef]
- Sapudom, J.; Alatoom, A.; Mohamed, W.K.E.; Garcia-Sabaté, A.; McBain, I.; Nasser, R.A.; Teo, J.C.M. Dendritic cell immune potency on 2D and in 3D collagen matrices. Biomater. Sci. 2020, 8, 5106–5120. [Google Scholar] [CrossRef] [PubMed]
- Santini, S.M. A New Type I IFN-Mediated Pathway for the Rapid Differentiation of Monocytes into Highly Active Dendritic Cells. Stem Cells 2003, 21, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Qu, C.; Brinck-Jensen, N.S.; Zang, M.; Chen, K. Monocyte-derived dendritic cells: Targets as potent antigen-presenting cells for the design of vaccines against infectious diseases. Int. J. Infect. Dis. 2014, 19, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tackett, N.; Bradley, J.H.; Moore, E.K.; Baker, S.H.; Minter, S.L.; DiGiacinto, B.; Arnold, J.P.; Gregg, R.K. Prolonged exposure to simulated microgravity diminishes dendritic cell immunogenicity. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Low, E.K.; Brudvik, E.; Kuhlman, B.; Wilson, P.F.; Almeida-Porada, G.; Porada, C.D. Microgravity Impairs DNA Damage Repair in Human Hematopoietic Stem/Progenitor Cells and Inhibits Their Differentiation into Dendritic Cells. Stem Cells Dev. 2018, 27, 1257–1267. [Google Scholar] [CrossRef]
- Seder, R.A.; Ahmed, R. Similarities and differences in CD4+ and CD8+ effector and memory T cell generation. Nat. Immunol. 2003, 4, 835–842. [Google Scholar] [CrossRef]
- Weyand, C.M.; Goronzy, J.J. The immunology of rheumatoid arthritis. Nat. Immunol. 2021, 22, 10–18. [Google Scholar] [CrossRef]
- Geenen, V. The thymus and the science of self. Semin. Immunopathol. 2021, 43, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Gensous, N.; Boizard-Moracchini, A.; Lazaro, E.; Richez, C.; Blanco, P. Update on the cellular pathogenesis of lupus. Curr. Opin. Rheumatol. 2021, 33, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Comi, G.; Bar-Or, A.; Lassmann, H.; Uccelli, A.; Hartung, H.P.; Montalban, X.; Sørensen, P.S.; Hohlfeld, R.; Hauser, S.L. Role of B Cells in Multiple Sclerosis and Related Disorders. Ann. Neurol. 2021, 89, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Ran, Z.; Yue-Bei, L.; Qiu-Ming, Z.; Huan, Y. Regulatory B Cells and Its Role in Central Nervous System Inflammatory Demyelinating Diseases. Front. Immunol. 2020, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Cogoli, A.; Tschopp, A.; Fuchs-Bislin, P. Cell sensitivity to gravity. Science 1984, 225, 228–230. [Google Scholar] [CrossRef]
- Schwarzenberg, M.; Pippia, P.; Meloni, M.A.; Cossu, G.; Cogoli-Greuter, M.; Cogoli, A. Signal transduction in T lymphocytes—A comparison of the data from space, the free fall machine and the random positioning machine. Adv. Sp. Res. 1999, 24, 793–800. [Google Scholar] [CrossRef]
- Hashemi, B.B.; Penkala, J.E.; Vens, C.; Huls, H.; Cubbage, M.; Sams, C.F. T cell activation responses are differentially regulated during clinorotation and in spaceflight. FASEB J. 1999, 13, 2071–2082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, D.; Pellis, N.R. Suppressed PHA activation of T lymphocytes in simulated microgravity is restored by direct activation of protein kinase C. J. Leukoc. Biol. 1998, 63, 550–562. [Google Scholar] [CrossRef]
- Simons, D.M.; Gardner, E.M.; Lelkes, P.I. Sub-mitogenic phorbol myristate acetate co-stimulation rescues the PHA-induced activation of both naïve and memory T cells cultured in the rotating-wall vessel bioreactor. Cell Biol. Int. 2009, 33, 882–886. [Google Scholar] [CrossRef]
- Simons, D.M.; Gardner, E.M.; Lelkes, P.I. Intact T cell receptor signaling by CD4+ T cells cultured in the rotating wall-vessel bioreactor. J. Cell. Biochem. 2010, 109, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Thiel, C.S.; Paulsen, K.; Bradacs, G.; Lust, K.; Tauber, S.; Dumrese, C.; Hilliger, A.; Schoppmann, K.; Biskup, J.; Golz, N.; et al. Rapid alterations of cell cycle control proteins in human T lymphocytes in microgravity. Cell Commun. Signal. 2012, 10, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, T.T.; Walther, I.; Li, C.-F.; Boonyaratanakornkit, J.; Galleri, G.; Meloni, M.A.; Pippia, P.; Cogoli, A.; Hughes-Fulford, M. The Rel/NF-κB pathway and transcription of immediate early genes in T cell activation are inhibited by microgravity. J. Leukoc. Biol. 2012, 92, 1133–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tauber, S.; Hauschild, S.; Crescio, C.; Secchi, C.; Paulsen, K.; Pantaleo, A.; Saba, A.; Buttron, I.; Thiel, C.S.; Cogoli, A.; et al. Signal transduction in primary human T lymphocytes in altered gravity—Results of the MASER-12 suborbital space flight mission. Cell Commun. Signal. 2013, 11, 32. [Google Scholar] [CrossRef] [Green Version]
- Tauber, S.; Hauschild, S.; Paulsen, K.; Gutewort, A.; Raig, C.; Hürlimann, E.; Biskup, J.; Philpot, C.; Lier, H.; Engelmann, F.; et al. Signal transduction in primary human T lymphocytes in altered gravity during parabolic flight and clinostat experiments. Cell. Physiol. Biochem. 2015, 35, 1034–1051. [Google Scholar] [CrossRef]
- Luo, H.; Wang, C.; Feng, M.; Zhao, Y. Microgravity inhibits resting T cell immunity in an exposure time-dependent manner. Int. J. Med. Sci. 2013, 11, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Kumarswamy, R.; Volkmann, I.; Thum, T. Regulation and function of miRNA-21 in health and disease. RNA Biol. 2011, 8, 706–713. [Google Scholar] [CrossRef] [Green Version]
- Hughes-Fulford, M.; Chang, T.T.; Martinez, E.M.; Li, C.F. Spaceflight alters expression of microRNA during T-cell activation. FASEB J. 2015, 29, 4893–4900. [Google Scholar] [CrossRef] [Green Version]
- Thiel, C.S.; Hauschild, S.; Huge, A.; Tauber, S.; Lauber, B.A.; Polzer, J.; Paulsen, K.; Lier, H.; Engelmann, F.; Schmitz, B.; et al. Dynamic gene expression response to altered gravity in human T cells. Sci. Rep. 2017, 7, 5204. [Google Scholar] [CrossRef] [Green Version]
- Thiel, C.S.; Huge, A.; Hauschild, S.; Tauber, S.; Lauber, B.A.; Polzer, J.; Paulsen, K.; Lier, H.; Engelmann, F.; Schmitz, B.; et al. Stability of gene expression in human T cells in different gravity environments is clustered in chromosomal region 11p15.4. npj Microgravity 2017, 3, 22. [Google Scholar] [CrossRef] [Green Version]
- Thiel, C.S.; Christoffel, S.; Tauber, S.; Vahlensieck, C.; de Zélicourt, D.; Layer, L.E.; Lauber, B.; Polzer, J.; Ullrich, O. Rapid cellular perception of gravitational forces in human Jurkat T cells and transduction into gene expression regulation. Int. J. Mol. Sci. 2020, 21, 514. [Google Scholar] [CrossRef] [Green Version]
- Martinez, E.M.; Yoshida, M.C.; Candelario, T.L.T.; Hughes-Fulford, M. Spaceflight and simulated microgravity cause a significant reduction of key gene expression in early T-cell activation. Am. J. Physiol. Integr. Comp. Physiol. 2015, 308, R480–R488. [Google Scholar] [CrossRef] [Green Version]
- Leung, S.; Smith, D.; Myc, A.; Morry, J.; Baker, J.R. OT-II TCR transgenic mice fail to produce anti-ovalbumin antibodies upon vaccination. Cell. Immunol. 2013, 282, 79–84. [Google Scholar] [CrossRef]
- Chang, T.T.; Spurlock, S.M.; Candelario, T.L.T.; Grenon, S.M.; Hughes-Fulford, M. Spaceflight impairs antigen-specific tolerance induction in vivo and increases inflammatory cytokines. FASEB J. 2015, 29, 4122–4132. [Google Scholar] [CrossRef] [Green Version]
- Bradley, J.H.; Stein, R.; Randolph, B.; Molina, E.; Arnold, J.P.; Gregg, R.K. T cell resistance to activation by dendritic cells requires long-term culture in simulated microgravity. Life Sci. Sp. Res. 2017, 15, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Bradley, J.H.; Barwick, S.; Horn, G.Q.; Ullrich, E.; Best, B.; Arnold, J.P.; Gregg, R.K. Simulated microgravity-mediated reversion of murine lymphoma immune evasion. Sci. Rep. 2019, 9, 14623. [Google Scholar] [CrossRef] [Green Version]
- Morabito, C.; Lanuti, P.; Caprara, G.A.; Marchisio, M.; Bizzarri, M.; Guarnieri, S.; Mariggiò, M.A. Physiological responses of jurkat lymphocytes to simulated microgravity conditions. Int. J. Mol. Sci. 2019, 20, 1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, J.; Thiel, C.S.; Tauber, S.; Stockmann, C.; Gassmann, M.; Ullrich, O. Expression of hypoxia-inducible factor 1α (Hif-1α) and genes of related pathways in altered gravity. Int. J. Mol. Sci. 2019, 20, 436. [Google Scholar] [CrossRef] [Green Version]
- Spielmann, G.; Agha, N.; Kunz, H.; Simpson, R.J.; Crucian, B.; Mehta, S.; Laughlin, M.; Campbell, J. B cell homeostasis is maintained during long-duration spaceflight. J. Appl. Physiol. 2019, 126, 469–476. [Google Scholar] [CrossRef]
- Tascher, G.; Gerbaix, M.; Maes, P.; Chazarin, B.; Ghislin, S.; Antropova, E.; Vassilieva, G.; Ouzren-Zarhloul, N.; Gauquelin-Koch, G.; Vico, L.; et al. Analysis of femurs from mice embarked on board BION-M1 biosatellite reveals a decrease in immune cell development, including B cells, after 1 wk of recovery on Earth. FASEB J. 2019, 33, 3772–3783. [Google Scholar] [CrossRef] [PubMed]
- Dang, B.; Yang, Y.; Zhang, E.; Li, W.; Mi, X.; Meng, Y.; Yan, S.; Wang, Z.; Wei, W.; Shao, C.; et al. Simulated microgravity increases heavy ion radiation-induced apoptosis in human B lymphoblasts. Life Sci. 2014, 97, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Martinac, B. Mechanosensitive ion channels: Molecules of mechanotransduction. J. Cell Sci. 2004, 117, 2449–2460. [Google Scholar] [CrossRef] [Green Version]
- Zhong, M.; Komarova, Y.; Rehman, J.; Malik, A.B. Mechanosensing Piezo channels in tissue homeostasis including their role in lungs. Pulm. Circ. 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Kuebler, W.M. Mechanotransduction by TRP Channels: General Concepts and Specific Role in the Vasculature. Cell Biochem. Biophys. 2010, 56, 1. [Google Scholar] [CrossRef]
- Wang, N. Review of Cellular Mechanotransduction. J. Phys. D Appl. Phys. 2017, 50, 233002. [Google Scholar] [CrossRef]
- Johnson, W.A. Two views of the same stimulus. Elife 2017, 6, e30191. [Google Scholar] [CrossRef] [Green Version]
- Martino, F.; Perestrelo, A.R.; Vinarský, V.; Pagliari, S.; Forte, G. Cellular Mechanotransduction: From Tension to Function. Front. Physiol. 2018, 9, 824. [Google Scholar] [CrossRef]
- Li, X.; Wang, J. Mechanical tumor microenvironment and transduction: Cytoskeleton mediates cancer cell invasion and metastasis. Int. J. Biol. Sci. 2020, 16, 2014–2028. [Google Scholar] [CrossRef]
- Jahed, Z.; Shams, H.; Mehrbod, M.; Mofrad, M.R.K. Mechanotransduction Pathways Linking the Extracellular Matrix to the Nucleus. Int. Rev. Cell. Mol. Biol. 2014, 310, 171–220. [Google Scholar] [CrossRef]
- Huang, H.; Kamm, R.D.; Lee, R.T. Cell mechanics and mechanotransduction: Pathways, probes, and physiology. Am. J. Physiol. Physiol. 2004, 287, C1–C11. [Google Scholar] [CrossRef] [PubMed]
- Das, T.; Safferling, K.; Rausch, S.; Grabe, N.; Boehm, H.; Spatz, J.P. A molecular mechanotransduction pathway regulates collective migration of epithelial cells. Nat. Cell Biol. 2015, 17, 276–287. [Google Scholar] [CrossRef]
- Peterson, J.R.; Mitchison, T.J. Small Molecules, Big Impact. Chem. Biol. 2002, 9, 1275–1285. [Google Scholar] [CrossRef] [Green Version]
- Oertel, A. Involvement of myosin in intracellular motility and cytomorphogenesis in Micrasterias. Cell Biol. Int. 2003, 27, 977–986. [Google Scholar] [CrossRef]
- Ji, Y.; Li, J.; Wei, Y.; Gao, W.; Fu, X.; Wang, Y. Substrate stiffness affects the immunosuppressive and trophic function of hMSCs via modulating cytoskeletal polymerization and tension. Biomater. Sci. 2019, 7, 5292–5300. [Google Scholar] [CrossRef]
- ElGindi, M.; Ibrahim, I.H.; Sapudom, J.; Garcia-Sabate, A.; Teo, J.C.M. Engineered Microvessel for Cell Culture in Simulated Microgravity. Int. J. Mol. Sci. 2021, 22, 6331. [Google Scholar] [CrossRef] [PubMed]
- Shanti, A.; Samara, B.; Abdullah, A.; Hallfors, N.; Accoto, D.; Sapudom, J.; Alatoom, A.; Teo, J.; Danti, S.; Stefanini, C. Multi-Compartment 3D-Cultured Organ-on-a-Chip: Towards a Biomimetic Lymph Node for Drug Development. Pharmaceutics 2020, 12, 464. [Google Scholar] [CrossRef]
- Court, M.; Malier, M.; Millet, A. 3D type I collagen environment leads up to a reassessment of the classification of human macrophage polarizations. Biomaterials 2019, 208, 98–109. [Google Scholar] [CrossRef]
- Garcia-Sabaté, A.; Mohamed, W.K.E.; Sapudom, J.; Alatoom, A.; Al Safadi, L.; Teo, J.C.M. Biomimetic 3D Models for Investigating the Role of Monocytes and Macrophages in Atherosclerosis. Bioengineering 2020, 7, 113. [Google Scholar] [CrossRef]
- Logie, C.; van Schaik, T.; Pompe, T.; Pietsch, K. Fibronectin-functionalization of 3D collagen networks supports immune tolerance and inflammation suppression in human monocyte-derived macrophages. Biomaterials 2021, 268, 120498. [Google Scholar] [CrossRef]
- Baker, B.M.; Chen, C.S. Deconstructing the third dimension—How 3D culture microenvironments alter cellular cues. J. Cell Sci. 2012, 125, 3015–3024. [Google Scholar] [CrossRef] [Green Version]
- Duval, K.; Grover, H.; Han, L.-H.; Mou, Y.; Pegoraro, A.F.; Fredberg, J.; Chen, Z. Modeling Physiological Events in 2D vs. 3D Cell Culture. Physiology 2017, 32, 266–277. [Google Scholar] [CrossRef]
- Sapudom, J.; Nguyen, K.-T.; Martin, S.; Wippold, T.; Möller, S.; Schnabelrauch, M.; Anderegg, U.; Pompe, T. Biomimetic tissue models reveal the role of hyaluronan in melanoma proliferation and invasion. Biomater. Sci. 2020, 8, 1405–1417. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.Y.; Yang, Z.; Han, B. Switch of macrophage fusion competency by 3D matrices. Sci. Rep. 2020, 10, 10348. [Google Scholar] [CrossRef]
- Sapudom, J.; Rubner, S.; Martin, S.; Pompe, T. Mimicking Tissue Boundaries by Sharp Multiparameter Matrix Interfaces. Adv. Healthc. Mater. 2016, 5, 1861–1867. [Google Scholar] [CrossRef]
- Galarza, S.; Kim, H.; Atay, N.; Peyton, S.R.; Munson, J.M. 2D or 3D? How cell motility measurements are conserved across dimensions in vitro and translate in vivo. Bioeng. Transl. Med. 2020, 5, e10148. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekaran, A.; Avci, H.X.; Ochalek, A.; Rösingh, L.N.; Molnár, K.; László, L.; Bellák, T.; Téglási, A.; Pesti, K.; Mike, A.; et al. Comparison of 2D and 3D neural induction methods for the generation of neural progenitor cells from human induced pluripotent stem cells. Stem Cell Res. 2017, 25, 139–151. [Google Scholar] [CrossRef]
- Sapudom, J.; Pompe, T. Biomimetic tumor microenvironments based on collagen matrices. Biomater. Sci. 2018, 6, 2009–2024. [Google Scholar] [CrossRef]
- Jensen, C.; Teng, Y. Is It Time to Start Transitioning From 2D to 3D Cell Culture? Front. Mol. Biosci. 2020, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Ravi, M.; Paramesh, V.; Kaviya, S.R.; Anuradha, E.; Solomon, F.D.P. 3D Cell Culture Systems: Advantages and Applications. J. Cell. Physiol. 2015, 230, 16–26. [Google Scholar] [CrossRef]
- Nicolas, J.; Magli, S.; Rabbachin, L.; Sampaolesi, S.; Nicotra, F.; Russo, L. 3D Extracellular Matrix Mimics: Fundamental Concepts and Role of Materials Chemistry to Influence Stem Cell Fate. Biomacromolecules 2020, 21, 1968–1994. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Kuga, H.; Morisaki, T.; Baba, E.; Sato, N.; Mizumoto, K.; Sueishi, K.; Tanaka, M.; Katano, M. Simulated Microgravity Culture System for a 3-D Carcinoma Tissue Model. Biotechniques 2002, 33, 1068–1076. [Google Scholar] [CrossRef]
- Tang, Y.; Xu, Y.; Xiao, Z.; Zhao, Y.; Li, J.; Han, S.; Chen, L.; Dai, B.; Wang, L.; Chen, B.; et al. The combination of three-dimensional and rotary cell culture system promotes the proliferation and maintains the differentiation potential of rat BMSCs. Sci. Rep. 2017, 7, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; Tian, W.; Yan, H.; Yue, L.; Zhang, Y.; Han, F.; Chen, X.; Li, Y. Rotary culture promotes the proliferation of MCF-7 cells encapsulated in three-dimensional collagen–alginate hydrogels via activation of the ERK1/2-MAPK pathway. Biomed. Mater. 2012, 7, 015003. [Google Scholar] [CrossRef]
- Ansorge, M.; Sapudom, J.; Chkolnikov, M.; Wilde, M.; Anderegg, U.; Möller, S.; Schnabelrauch, M.; Pompe, T. Mimicking Paracrine TGFβ1 Signals during Myofibroblast Differentiation in 3D Collagen Networks. Sci. Rep. 2017, 7, 5664. [Google Scholar] [CrossRef] [PubMed]
- Sapudom, J.; Wu, X.; Chkolnikov, M.; Ansorge, M.; Anderegg, U.; Pompe, T. Fibroblast fate regulation by time dependent TGF-β1 and IL-10 stimulation in biomimetic 3D matrices. Biomater. Sci. 2017, 5, 1858–1867. [Google Scholar] [CrossRef] [PubMed]
- Hallfors, N.; Shanti, A.; Sapudom, J.; Teo, J.; Petroianu, G.; Lee, S.; Planelles, L.; Stefanini, C. Multi-Compartment Lymph-Node-on-a-Chip Enables Measurement of Immune Cell Motility in Response to Drugs. Bioengineering 2021, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Witkowski, M.T.; Harris, J.; Dolgalev, I.; Sreeram, S.; Qian, W.; Tong, J.; Chen, X.; Aifantis, I.; Chen, W. Leukemia-on-a-chip: Dissecting the chemoresistance mechanisms in B cell acute lymphoblastic leukemia bone marrow niche. Sci. Adv. 2020, 6, eaba5536. [Google Scholar] [CrossRef]
- Yuan, M.; Liu, H.; Zhou, S.; Zhou, X.; Huang, Y.; Hou, F.; Jiang, W. Integrative Analysis of Regulatory Module Reveals Associations of Microgravity with Dysfunctions of Multi-body Systems and Tumorigenesis. Int. J. Mol. Sci. 2020, 21, 7585. [Google Scholar] [CrossRef] [PubMed]
- Nassef, M.Z.; Melnik, D.; Kopp, S.; Sahana, J.; Infanger, M.; Lützenberg, R.; Relja, B.; Wehland, M.; Grimm, D.; Krüger, M. Breast Cancer Cells in Microgravity: New Aspects for Cancer Research. Int. J. Mol. Sci. 2020, 21, 7345. [Google Scholar] [CrossRef]
- Fukazawa, T.; Tanimoto, K.; Shrestha, L.; Imura, T.; Takahashi, S.; Sueda, T.; Hirohashi, N.; Hiyama, E.; Yuge, L. Simulated microgravity enhances CDDP-induced apoptosis signal via p53-independent mechanisms in cancer cells. PLoS ONE 2019, 14, e0219363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braddock, M. From Target Identification to Drug Development in Space: Using the Microgravity Assist. Curr. Drug Discov. Technol. 2020, 17, 45–56. [Google Scholar] [CrossRef]
- Schmidt, M.A.; Schmidt, C.M.; Hubbard, R.M.; Mason, C.E. Why Personalized Medicine Is the Frontier of Medicine and Performance for Humans in Space. New Space 2020, 8, 63–76. [Google Scholar] [CrossRef]
- Sapudom, J.; Waschke, J.; Franke, K.; Hlawitschka, M.; Pompe, T. Quantitative label-free single cell tracking in 3D biomimetic matrices. Sci. Rep. 2017, 7, 14135. [Google Scholar] [CrossRef] [Green Version]
- Scherf, N.; Franke, K.; Glauche, I.; Kurth, I.; Bornhäuser, M.; Werner, C.; Pompe, T.; Roeder, I. On the symmetry of siblings: Automated single-cell tracking to quantify the behavior of hematopoietic stem cells in a biomimetic setup. Exp. Hematol. 2012, 40, 119–130.e9. [Google Scholar] [CrossRef]
- Pearson, Y.E.; Lund, A.W.; Lin, A.W.H.; Ng, C.P.; Alsuwaidi, A.; Azzeh, S.; Gater, D.L.; Teo, J.C.M. Non-invasive single-cell biomechanical analysis using live-imaging datasets. J. Cell Sci. 2016, 129, 3351–3364. [Google Scholar] [CrossRef] [Green Version]
- Kräter, M.; Sapudom, J.; Bilz, N.; Pompe, T.; Guck, J.; Claus, C. Alterations in Cell Mechanics by Actin Cytoskeletal Changes Correlate with Strain-Specific Rubella Virus Phenotypes for Cell Migration and Induction of Apoptosis. Cells 2018, 7, 136. [Google Scholar] [CrossRef] [Green Version]
- Lühr, J.J.; Alex, N.; Amon, L.; Kräter, M.; Kubánková, M.; Sezgin, E.; Lehmann, C.H.K.; Heger, L.; Heidkamp, G.F.; Smith, A.-S.; et al. Maturation of Monocyte-Derived DCs Leads to Increased Cellular Stiffness, Higher Membrane Fluidity, and Changed Lipid Composition. Front. Immunol. 2020, 11, 2908. [Google Scholar] [CrossRef] [PubMed]
- Menachery, A.; Sapudom, J.; Vembadi, A.; Alatoom, A.; Teo, J.; Qasaimeh, M.A. Dielectrophoretic characterization of dendritic cell deformability upon maturation. Biotechniques 2021, 70, 29–36. [Google Scholar] [CrossRef]
- Alatoom, A.; Sapudom, J.; Soni, P.; Mohamed, W.K.E.; Garcia-Sabaté, A.; Teo, J. Artificial Biosystem for Modulation of Interactions between Antigen-Presenting Cells and T Cells. Adv. Biosyst. 2020, 4, 2000039. [Google Scholar] [CrossRef]
- Blumenthal, D.; Chandra, V.; Avery, L.; Burkhardt, J.K. Mouse T cell priming is enhanced by maturation-dependent stiffening of the dendritic cell cortex. Elife 2020, 9, e55995. [Google Scholar] [CrossRef] [PubMed]
- Corydon, T.J.; Kopp, S.; Wehland, M.; Braun, M.; Schütte, A.; Mayer, T.; Hülsing, T.; Oltmann, H.; Schmitz, B.; Hemmersbach, R.; et al. Alterations of the cytoskeleton in human cells in space proved by life-cell imaging. Sci. Rep. 2016, 6, 20043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuest, S.L.; Richard, S.; Kopp, S.; Grimm, D.; Egli, M. Simulated Microgravity: Critical Review on the Use of Random Positioning Machines for Mammalian Cell Culture. Biomed Res. Int. 2015, 2015, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neelam, S.; Lee, A.; Lane, M.A.; Udave, C.; Levine, H.G.; Zhang, Y. Module to Support Real-Time Microscopic Imaging of Living Organisms on Ground-Based Microgravity Analogs. Appl. Sci. 2021, 11, 3122. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
ElGindi, M.; Sapudom, J.; Ibrahim, I.H.; Al-Sayegh, M.; Chen, W.; Garcia-Sabaté, A.; Teo, J.C.M. May the Force Be with You (Or Not): The Immune System under Microgravity. Cells 2021, 10, 1941. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10081941
ElGindi M, Sapudom J, Ibrahim IH, Al-Sayegh M, Chen W, Garcia-Sabaté A, Teo JCM. May the Force Be with You (Or Not): The Immune System under Microgravity. Cells. 2021; 10(8):1941. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10081941
Chicago/Turabian StyleElGindi, Mei, Jiranuwat Sapudom, Ibrahim Hamed Ibrahim, Mohamed Al-Sayegh, Weiqiang Chen, Anna Garcia-Sabaté, and Jeremy C. M. Teo. 2021. "May the Force Be with You (Or Not): The Immune System under Microgravity" Cells 10, no. 8: 1941. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10081941